Circovirus et pathologies associées chez les animaux

revue
Circovirus et pathologies associées
chez les animaux
B. Grasland
L. Bigarré
P. Blanchard
C. Loizel
Y. Blanchard
C. de Boisséson
A. Jestin
Unité de génétique virale et biosécurité,
Agence française de sécurité sanitaire
des aliments (Afssa),
BP53,
22440 Ploufragan
Résumé.Les Circovirus représentent un des deux genres de la famille des
Circoviridae. À ce jour, seuls les suidés (porc et sanglier) et les oiseaux ont été
identifiés comme leurs hôtes. Ils possèdent tous un génome à ADN circulaire
simple brin d’environ 2 000 nucléotides. Leur ADN contient une séquence no-
nanucléotidique nécessaire pour la réplication, flanquée de séquences répétées
inversées représentant un motif tige-boucle et deux cadres de lecture principaux
d’orientation opposée, l’un codant pour la protéine Rep associée à la réplication
et l’autre pour la protéine de capside Cap. Tous les circovirus décrits à ce jour,
excepté le circovirus porcin de type 1 (PCV1), sont associés à des pathologies
caractérisées par une immunosuppression ou immunodépression de leur hôte.
Des lésions histopathologiques, telles que les inclusions intracytoplasmiques
dans les macrophages et des déplétions lymphocytaires des lymphocytes T et B
au niveau des organes lymphoïdes, sont généralement constatées.Actuellement,
aucun moyen de prophylaxie médicale n’est disponible pour prévenir les infec-
tions à circovirus.
Mots clés :circovirus, ADN simple brin, génome circulaire, cofacteur
d’infection
Abstract.Animal circoviruses belong to the Circovirus genus of the Circoviri-
dae family. Nowadays, only swine and birds were identified as circovirus hosts.
Circoviruses have a single-stranded circular genome of approximately 2000
nucleotide long. DNA of these viruses possesses : (i) a nonanucleotide sequence
essential for replication, flanked by inverted repeat sequences, a palindrome that
has the potential to form a stem-loop structure and (ii) two major ORFs, located
on the viral and complementary strands, which encode respectively the
replication-associated protein (Rep) and the capsid protein (Cap). All the cir-
coviruses described at the present time, except porcine circovirus of type 1, are
associated with immunosuppressive or immunodepressive diseases. Histopatho-
logical lesions such as cytoplasmic inclusions of virus in histiocytic cells and T
and B lymphocyte depletion in lymphoid organs, are commonly noticed. No
medical prophylaxis of circovirus infections is currently available.
Key words:circovirus, single-stranded DNA, circular genome, infection
cofactor
Historique et classification
Les circovirus infectant les vertébrés possèdent un ADN
circulaire simple brin et présentent des similitudes avec les
virus appartenant aux Geminiviridae et aux Nanoviridae
qui infectent les plantes. Le génome des Geminiviridae est
constitué d’une ou deux molécules d’ADN simple brin
circulaire de 2 500 à 3 000 nucléotides (tableau 1) [1]. Ce-
lui des Nanoviridae est réparti sur au moins6à8molécules
d’ADN simple brin circulaire, d’environ 1 000 nucléotides
chacune [2]. Les particules virales de ces deux familles sont
non enveloppées, de forme icosaédrique simple d’un dia-
mètre de 18 nm pour les nanovirus et de forme icosaédrique
double d’environ 18 × 30 nm pour les géminivirus (ta-
Virologie 2005, 9 : 431-42
Virologie, Vol. 9, n° 6, novembre-décembre 2005
431
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

bleau 1). Il a été émis l’hypothèse que les circovirus déri-
veraient d’un nanovirus qui aurait recombiné avec un virus
infectant les vertébrés [3].
En 1974, des particules virales ressemblant à des picorna-
virus sont observées comme contaminant d’une lignée cel-
lulaire de reins de porc (PK15) [4]. Il faudra attendre 1982
pour réaliser que ces particules virales contenaient un gé-
nome à ADN simple brin, circulaire, de petite taille, à
l’origine du nom de circovirus porcin (PCV) [5]. Par la
suite, des virus de même structure génomique, le virus de la
maladie du bec et du plumage des Psittacidae (beak and
feather disease virus, BFDV) et le virus de l’anémie du
poulet (chicken anemia virus, CAV), sont découverts chez
les oiseaux. De par leurs caractéristiques biochimiques et
morphologiques communes [6, 7], ces trois virus ont été
regroupés dans la famille des Circoviridae, créée en 1995
par l’International Committee for the Taxonomy of Viruses
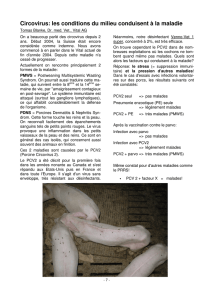
(ICTV) [8]. En 1996, une nouvelle pathologie appelée
maladie d’amaigrissement du porcelet (MAP) fait son ap-
parition en France et en Espagne [9]. Elle affecte sévère-
ment les rendements des élevages porcins, la mortalité
pouvant atteindre 20 %, voire plus dans les cas les plus
sévères. Elle touche les porcelets en croissance âgés de 2 à
4 mois et est aujourd’hui présente dans la majorité des pays
producteurs de porcs. Un circovirus a été identifié à partir
de lésions de porcelets malades. Il s’est révélé être différent
du circovirus porcin identifié dès 1982 [10], en termes
d’antigénicité et de séquence nucléotidique. Le circovirus
porcin isolé de cellules PK15 est alors appelé circovirus
porcin de type 1 (porcine circovirus type 1, PCV1) et celui
isolé d’animaux atteints de MAP, circovirus porcin de
type 2 (porcine circovirus type 2, PCV2). En 2005, l’ICTV
distingue deux genres dans la famille des Circoviridae :le
genre Gyrovirus avec un représentant unique, le CAV, et le
genre Circovirus incluant les deux autres virus déjà connus
et de nouveaux membres découverts chez des espèces
aviaires dont le circovirus du pigeon (pigeon circovirus,
PiCV), de l’oie (goose circovirus, GoCV) et du canari
(canary circovirus, CaCV) (tableau 1) [11]. Des espèces
isolées chez le pinson (finch circovirus, FiCV), le canard
(duck circovirus, DuCV) et la mouette (gull circovirus,
GuCV) sont en attente d’y être classées [11]. Le Torque
teno virus (TTV) et le TTV-like mini-virus (TLMV), décou-
verts chez l’homme à partir de 1997 et décrits initialement
comme « circovirus humains », sont désormais regroupés
dans le genre Anellovirus, dans l’attente d’être assignés à
une famille [12].
La revue présente les circovirus, tels que définis dans la
dernière classification présentée par l’ICTV, et leurs patho-
logies associées. Les auteurs s’appuient essentiellement sur
les modèles des circovirus porcins qui ont été très étudiés
ces dernières années.
Les virus du genre circovirus
Caractéristiques biologiques
Spectre d’hôte
Le PCV1 a été isolé dans la lignée cellulaire de reins de
porc PK15 (ATCC-CCL33) [4]. Il a été montré qu’il pou-
vait infecter et se multiplier dans d’autres lignées cellulai-
res porcines primaires ou semi-continues [5, 13]. Des étu-
des sérologiques ont démontré que des anticorps contre le
PCV1 étaient communément présents chez les porcs dans
de nombreux pays [14]. Cependant, des réactions croisées
avec le PCV2 pourraient avoir conduit à une surestimation
Tableau 1.Classification et caractéristiques générales des familles Nanoviridae,Geminiviridae et Circoviridae
Famille Nanoviridae Geminiviridae Circoviridae
Genre
Gyrovirus
Genre
Circovirus
Chicken anemia virus (CAV) Beak and feather disease
virus (BFDV)
Canary circovirus (CaCV)
Goose circovirus (GoCV)
Pigeon circovirus (PiCV)
Porcine circovirus 1 (PCV1)
Porcine circovirus 2 (PCV2)
Hôte Plantes Plantes Vertébrés Vertébrés
Génome ADN circulaire simple brin ADN circulaire simple brin ADN circulaire simple brin Vertébrés
6à8segments
de1kb
1 ou 2 segments
de2,5à3kb
1 segment de 1,8à2kb
Virion Non enveloppé Non enveloppé Non enveloppé Non enveloppé
capsidede18à
20 nm de Ø*
capside géminée
de 20 nm de Ø*
capsidede14à20nmdeØ*
*Ø diamètre.
revue
Virologie, Vol. 9, n° 6, novembre-décembre 2005
432
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

de la prévalence du PCV1. Le PCV2 a quant à lui été isolé
chez des suidés (porc et sanglier) en Europe [15, 16], en
Amérique du Nord, centrale et du Sud [16, 17] ainsi qu’en
Asie [18]. Des anticorps anti-PCV2 ont été détectés dans
des sera antérieurs de 30 ans à l’émergence de la MAP,
indiquant que les porcs étaient infectés par le PCV2 bien
avant l’isolement du virus [19]. Les circovirus porcins
n’ont été retrouvés que chez les suidés, indiquant une
grande spécificité d’hôte. Les recherches sérologiques ef-
fectuées dans les populations humaines exposées et anima-
les (bovins, ovins, poulet, lapin) sont restées négatives [20,
21]. Des essais d’inoculation de PCV1 et PCV2 sur diver-
ses lignées bovines et ovines ont montré que les virus
pouvaient infecter ces cellules sans qu’il y ait cependant
production de virions [20]. En outre, l’infection de diffé-
rentes lignées cellulaires humaines et simiennes par le
PCV1 ou le PCV2 est non productive de virus bien que le
PCV1, comme le PCV2, puisse entrer et se répliquer dans
certaines cellules humaines [22].
Plus de 60 espèces d’oiseaux de la famille des Psittacidae,
comprenant notamment les perroquets et les perruches,
peuvent être infectées par des variants du BFDV [23]. Le
virus a une distribution mondiale et se rencontre dans les
populations sauvages d’Amérique du Sud, d’Afrique,
d’Australie et du Pacifique sud ainsi que dans les popula-
tions captives. À l’heure actuelle, chaque circovirus aviaire
n’a qu’un seul hôte connu, l’espèce chez laquelle il a été
détecté. Mis à part les circovirus porcins, les autres virus
n’ont pu être répliqués sur système cellulaire in vitro.
Morphologie
Les particules de circovirus sont non enveloppées, en forme
d’icosaèdre et ont un diamètre compris entre 14 et 20 nm
(figure 1). Les circovirus porcins et le BFDV seraient
constitués de 12 pentamères comprenant 60 sous-unités ré-
pétées d’une protéine de capside unique [24].
Propriétés physicochimiques et biologiques
La densité du PCV1 est évaluée à 1,37 g/ml en chlorure de
césium. Le virus ne provoque pas l’hémagglutination
d’érythrocytes de nombreuses espèces dont le porc. Il ré-
siste à l’inactivation à pH 3 ainsi qu’au chloroforme, et
reste stable à 56 et 70 °C pendant 15 minutes [20]. Peu de
données existent sur les propriétés physicochimiques du
PCV2. In vitro, ce virus est résistant aux désinfectants
contenant de la chlorhexidine, du formaldéhyde, de l’iodine
et des alcools. Cependant, il montre une sensibilité aux
désinfectants à base de phénol, d’ammonium quaternaire,
d’hydroxyde de sodium et d’agents oxydants [25].
Le BFDV agglutine les érythrocytes de perroquets mais pas
ceux de l’homme, du porc, de l’oie et du cobaye [26] même
si, pour ce dernier, les résultats sont contradictoires [27].
Génome et expression
Organisation génomique et réplication
Le génome des circovirus est une molécule d’ADN simple
brin circulaire, d’une taille d’environ 2 000 nucléotides
(figure 2). Leur structure est composée d’une région inter-
génique comprise entre deux cadres de lecture majeurs,
d’orientation opposée et appelés ORF1 et 2. L’ORF1, situé
sur le brin viral de polarité positive (+), est compris dans le
gène rep qui code pour deux protéines initiatrices de la
réplication (Rep et Rep’) tandis que l’ORF2, situé sur le
brin de polarité négative (–), est compris dans le gène cap
qui code pour l’unique protéine de structure de la capside.
L’origine de réplication se trouve dans la région intergéni-
que qui inclut une séquence nonanucléotidique flanquée de
séquences palindromiques, l’ensemble formant un motif
tige-boucle (figure 2). En aval de cette tige-boucle, se
trouvent des séquences hexanucléotidiques répétées. La
réplication du génome impliquerait un mécanisme en cer-
cle roulant comme pour les Geminiviridae. Cependant,
chez les circovirus, l’originalité repose sur le choix de la
matrice à copier au niveau de la tige-boucle, reposant sur
une organisation en melting-pot, différente de celle quali-
fiée de « cruciforme » utilisée par les Geminiviridae. Dans
ce modèle, une forme réplicative double brin est obtenue à
partir du génome viral simple brin après infection des
cellules. Puis un complexe REP, formé des protéines Rep et
Rep’, se lie à la séquence nonanucléotidique comprise dans
la séquence de la tige-boucle de l’intermédiaire bicaténaire
superenroulé. Le complexe déstabilise l’origine de réplica-
tion et coupe alors la séquence nonanucléotidique en amont
de l’avant-dernier nucléotide (A) (la position du nucléotide
1 du génome étant fixé sur ce nucléotide dans la figure 2), ce
qui engendre la création d’une extrémité 3′OH libre.
Aucune structure en croix n’est formée au niveau de l’ori-
100 nm
100 nm
Figure 1. Particules virales de circovirus porcin de type 2 semi-
purifiées après coloration négative au PTA (microscope électroni-
que × 200 000). Des particules de 20 nm de diamètre environ sont
visibles.
revue
Virologie, Vol. 9, n° 6, novembre-décembre 2005
433
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

gine de réplication, mais plutôt une sphère d’instabilité,
appelée melting-pot où les brins (+) et (–) sont proches mais
non liés par des liaisons hydrogène. Le brin (–) et le brin (+)
du génome viral au niveau de la séquence palindromique de
la tige-boucle sont donc disponibles simultanément comme
modèles pendant les phases d’induction et de terminaison
de la réplication [28].
Les circovirus dépendent de l’ADN polymérase de leur
hôte pour effectuer leur cycle de réplication du génome et
produire la forme réplicative double brin. La nécessité de
cette polymérase conforte le fait que les circovirus porcins
requièrent des cellules en phase S du cycle cellulaire pour
réaliser un cycle d’infection complet [29]. Des études in
vivo ont permis d’établir que les cellules supportant la
réplication du PCV2 dépendaient du stade de développe-
ment de leur hôte au moment de l’infection. Chez des fœtus
infectés in utero, le PCV2 a été détecté dans des cardio-
myocytes, des hépatocytes et des macrophages au début de
la gestation et dans des macrophages majoritairement en fin
de gestation [30]. Des études réalisées in vitro avec une
lignée cellulaire de monocytes ont montré que le PCV2
entre dans ces cellules par endocytose dépendante de la
clathrine essentiellement et que l’acidification du compar-
timent endosomal est nécessaire à l’infection par le PCV2
[31].
Le génome du PCV1 compte 1 759 nucléotides et celui du
PCV2, 1 767 ou 1 768, en fonction des souches [11, 15].
L’origine de réplication du PCV1 a pu être expérimentale-
ment bornée à un fragment de 111 bases (b) de longueur
englobant le nonanucléotide 5’TAGTATTAC ainsi que le
promoteur du gène rep [32]. La séquence de ce motif
nonanucléotidique ne varie que par la première base chez le
PCV2 5’AAGTATTAC. Des modifications de nucléotide
dans ce motif par mutagenèse dirigée montre que la sé-
quence importante pour la réplication peut être condensée à
un octanucléotide 5’AxTAxTAC [33]. En aval de la tige-
boucle, se trouvent quatre répétitions de l’hexamère 5’CG-
GCAG (figure 2) qui forment un site de fixation aux protéi-
nes codées par le gène rep. Les gènes rep des deux PCV
présentent plus de 85 % de similitude alors que les gènes
cap sont moins conservés avec 62 % de similitude.
La séquence nucléotidique du génome du BFDV est com-
posée de 1 993 nucléotides et comprend la même séquence
nonanucléotidique indispensable pour la réplication que le
PCV1. Deux répétitions d’un motif de 8 nucléotides
5’GGGGCACC sont situées en aval du motif tige-boucle.
Le circovirus du pigeon (PiCV) possède quant à lui un
génome de 2 036 ou 2 037 b, celui de l’oie (GoCV) de
1 821 b, du canard (DuCV) de 1 996 b et du canari (CaCV)
de 1 952 b [11].
Transcription et protéines virales
Les génomes des circovirus porcins présentent deux cadres
de lecture permettant, par un phénomène d’épissage alter-
natif, la synthèse de trois protéines majeures : les protéines
Rep et Rep’ via le brin d’ADN viral (+) et la protéine Cap
via le brin (–) complémentaire (figure 3). Un troisième
cadre de lecture, appelé ORF3, a été mis en évidence chez
le PCV2. Le gène serait situé au niveau du gène rep mais
transcrit en sens inverse. L’ARNm de 315 nt coderait pour
une protéine qui jouerait un rôle dans l’induction de l’apop-
tose dans les cellules infectées [34].
L’incapacité de propager les circovirus autres que les cir-
covirus porcins en culture cellulaire aura été un frein à
l’analyse des transcrits produits chez les autres circovirus.
Néanmoins, pour le BFDV, deux protéines majeures de 26
et 23 kDa associées à la capside ont été caractérisées à
partir de virus purifiés provenant de plumes infectées [35].
Leur fonction n’est cependant toujours pas établie.
–Cap. Pour les PCV, un ARNm majeur est transcrit à partir
du brin complémentaire. Il subit un épissage produisant un
transcrit codant pour la protéine Cap. Chez le PCV1, le site
d’induction de la transcription est localisé au nt 457, le site
d’épissage compris entre les nt 348 et 1724, et le site de
terminaison au nt 998 (figure 3). La région codant pour la
Génome des
circovirus
~ 2000 nt
ORF2
cap ORF1
rep
Tige boucle
GC
TA
CG
GC
CG
GC
TA
GC
AT
AT
G
13 19 30 36
5’ 3’
CGGCAG CGGCAG CACCT CGGCAG CGTCAG
C
T
G
T
AG T
A
T
C
T
A
1
Origine de
réplication
Figure 2. Organisation génomique générale des circovirus ani-
maux. Deux cadres de lecture majeurs, de sens contraire, sont
compris dans les gènes rep et cap. Une partie de la région
intergénique du PCV1 contenant le motif tige-boucle et la sé-
quence nonamérique essentielle pour la réplication (indiquée en
gras), est agrandie en exemple. La position du premier nucléotide
ainsi que celles des hexamères sont indiquées.
revue
Virologie, Vol. 9, n° 6, novembre-décembre 2005
434
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

protéine Cap s’étend du nt 1 723 au nt 1 024. Pour le PCV2,
les sites d’induction et de terminaison sont les nt 469 et
1 005. La région d’épissage est bornée par les nt 361 et
1 737. La région codante est comprise entre les nt 1 735 et
1 037 [36]. L’épissage se fait donc en dehors du cadre de
lecture. Chez ces deux virus, les ARNm codent pour une
protéine de 233 acides aminés. Les protéines Cap des PCV
présentent 67 % d’homologie. Leurs masses moléculaires
calculées sont de 28 kDa, valeur inférieure aux masses
observées après électrophorèse sur gel dénaturant
(30 kDa), ce qui suggère des modifications post-
traductionnelles [37]. La protéine Cap se localise dans le
noyau, le ciblage nucléaire faisant intervenir les 41 pre-
miers acides aminés de la région N-terminale. Une analyse
par mutagenèse dirigée confirme la présence de deux si-
gnaux de localisation nucléaire potentiels, constitués d’aci-
des aminés basiques comprenant les résidus aminés 12 à 18
et 34 à 41 [38]. La détection de la protéine n’est possible
que 24 heures après infection in vitro, suggérant que la
protéine interviendrait à la fin du cycle de multiplication.
Quatre épitopes immunodominants, situés au niveau des
acides aminés 65-87, 113-147, 157-183 et 193-207, ont été
identifiés dans la protéine de capside du PCV2 [39].
–Rep et Rep’. Chez le PCV1, un ARNm d’une taille de
978 nt a été détecté et code pour la protéine Rep (312 acides
aminés) impliquée dans la réplication (figure 3). Cet
Génome du PCV1
ARNm Cap
457 348 1724
1759/1
1723 1024
998
ARNm Cap
469 361 1737
1735 1037
1005
998 997
Génome du PCV2
1768/1
1005 1004
ARNm Rep et Rep’ ARNm Rep et Rep’
Rep
Rep’
19
19 403 787 997
997
47
47 403 787 934
983
Rep
Rep’
18
18 416 800 1004
1004
51
51 416 800 967
992
Figure 3. Génomes et transcrits principaux du PCV1 et du PCV2. La direction de transcription est indiquée par les flèches au niveau des
génomes. Les séquences codantes apparaissent en gris ainsi que leur localisation par rapport au génome, signalée sous chaque transcrit.
Les queues polyA de chaque ARNm et les sites d’épissage sont indiqués respectivement par et ....
revue
Virologie, Vol. 9, n° 6, novembre-décembre 2005
435
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%