Cours 5 : Méiose – Fécondation – Cycle biologique

Cours 5 : Méiose – Fécondation – Cycle biologique – Anomalies
chromosomiques
Introduction et rappel :
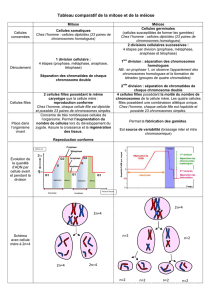
Chez les organismes eucaryotes, il existe deux types de divisions cellulaires : la mitose et la méiose.
La mitose concerne les cellules somatiques et assure la naissance de cellules identiques à la cellule mère lors de
la multiplication asexuée (elle conserve donc l'information génétique et assure la prolifération cellulaire). La mitose
intervient généralement sur des cellules diploïdes mais elle est aussi réalisée par des cellules haploïdes chez
certaines espèces (le champignon Sordaria macrospora, la laitue de mer…).
Les cellules somatiques possèdent deux exemplaires de chromosomes : les chromosomes identiques sont dits
homologues et sont organisés par paire ; la cellule est diploïde.
Les cellules sexuelles (germinales) ne présentent qu’un seul exemplaire de chaque chromosome : la cellule est
haploïde.
Différences entre mitose et méiose :
La méiose aboutit à la production de cellules sexuelles haploïdes (ou gamètes) pour la reproduction. Chez les
animaux, la méiose est un processus se déroulant durant la gamétogénèse (spermatogenèse ou ovogenèse), c'est-à-
dire durant l'élaboration des gamètes (les spermatozoïdes chez le mâle et les ovules chez la femelle) chez les
espèces diploïdes.
En plus du rôle de division, la méiose a un rôle dans le brassage génétique (mélange des gènes) et ce, grâce à
deux mécanismes de recombinaison génétique : le brassage inter-chromosomique et le brassage intra-
chromosomique.
La méiose se déroule en plusieurs étapes formant un ensemble de deux divisions cellulaires, successives et
inséparables. La première division méiotique est dite réductionnelle car elle permet de passer de 2n chromosomes
doubles à n chromosomes doubles (= séparation des chromosomes homologues). La seconde est dite équationnelle
car elle conserve le nombre de chromosomes : on passe de n chromosomes doubles à n chromosomes simples (=
séparation des chromatides). La méiose permet ainsi la formation de 4 cellules filles haploïdes (ou gamètes).
On commencera par décrire toutes les phases de la méiose et des brassages génétiques correspondants. Parce
que la prépondérance des phases haploïde et diploïde est variable entre les espèces, nous détaillerons ensuite
différents cas de cycle biologique (Partie 2). Dans une 3ème partie, on s’intéressera plus spécifiquement, aux
anomalies chromosomiques et pathologies associées.

Partie 1 : La méiose et la fécondation sont à la base des cycles de
reproduction sexuée
Deux animations pour illustrer la méiose :
- Une simple : http://www.biologieenflash.net/animation.php?ref=bio-0051-2
- Une complexe (qui montre les conséquences génétique de la méiose) :
http://www.snv.jussieu.fr/bmedia/Meiose/Meiose_fichiers/Meiose3.swf
I- Les étapes de la méiose et leur brassage génétique
a- Méiose 1 ou division réductionnelle
La méiose 1 est précédée par une interphase au cours de laquelle chacun des chromosomes se réplique et se
retrouve constitué de deux chromatides-sœurs, parfaites copies l'une de l'autre.
La première division de la méiose permet la réduction chromatique (réduction du nombre de chromosomes :
passage de 2n à n) par séparation des chromosomes homologues. Elle est constitué de 4 phases d'une division
cellulaire avec quelques caractéristiques en plus propre à la méiose 1.
La prophase 1 :
L'enveloppe nucléaire se fragmente et disparaît. Les chromosomes commencent à se condenser. Le fuseau
méiotique se forme. Les chromosomes homologues s'apparient (= s'associent) deux à deux pour former n tétrades
(ou bivalents) = complexes de 4 chromatides. Chaque chromosome bivalent contient un chromosome paternel et
maternel. C'est la phase la plus longue de la méiose. Les chromatides homologues se croisent en plusieurs endroits
et forment des chiasmata (points de chevauchement de deux chromatides). Il y a alors possibilité d'échanges de
fragments de chromatides entre chromatides homologues. Ce phénomène, appelé crossing over, est à l'origine du
brassage intrachromosomique se produisant en prophase 1 de méiose. Il permet une recombinaison des allèles

entre chromosomes homologues, c'est à dire une répartition nouvelle des allèles sur un chromosome. Les
chromatides sœurs ne portent plus les mêmes allèles. On parle de chromosomes recombinés, c'est à dire des
chromosomes possédant des informations maternelles et paternelles. Ce phénomène augmente la diversité des
gamètes produits dans le cas de génotype hétérozygote.
Exemple de crossing over (ou recombinaison homologue) entre deux chromosomes homologues maternel et paternel
A ne pas connaître : La prophase I est divisée en cinq étapes qui correspondent à cinq états caractéristiques de
la chromatine :
- leptotène : les chromosomes se condensent. Les télomères s’attachent à l’enveloppe nucléaire.
- zygotène : appariement des chromosomes homologues via un complexe synaptonémal (qui est composé de
protéines et enzymes, qui forment les éléments latéraux et l’élément central). L'appariement des
chromatides homologues démarre sur l'enveloppe nucléaire et progresse le long des chromosomes «
comme une fermeture éclair ». Il y a donc formation des bivalents (ou tétrades).
- pachytène : au niveau central du complexe synaptonémal, des protéines (qui constituent les nodules de
recombinaison) effectuent les enjambements (2-3/ chromosome).
- diplotène : disparition du complexe synaptonémal. Séparation des chromosomes homologues sauf au niveau
des chiasmata entre les chromatides. Décondensation et forte transcription.
- diacinèse : Recondensation de la chromatine. Détachement des télomères. A la fin, rupture de l’enveloppe
nucléaire.

le crossing-over (ou enjambement): échange réciproque d'une portion de chromatide entre deux chromosomes
homologues.
le chiasma (pluriel : chiasmata) : c’est l’entrecroisement observé de deux chromatides. C’est au niveau des
chiasmata que se font les crossing-over.
La métaphase 1 :
Les bivalents fixés aux microtubules kinétochoriens s'alignent sur le plan équatorial (formant la plaque
équatoriale). Les chiasmata (singulier : chiasma) sont alignés et les centromères de chaque bivalent se retrouvent de
part et d'autre du plan équatorial.
L'anaphase 1 :
Les chromosomes homologues, dirigés par les fibres du fuseau, se séparent. Chacun migre vers un pôle de la
cellule emportant ses deux chromatides sœurs. Il n'y a pas ici de division des centromères. Au cours de l'anaphase 1
se produit le brassage interchromosomique. Il est dû à la répartition aléatoire et indépendante des chromosomes, il
dépend du positionnement des chromosomes bivalents en métaphase 1. En effet, chaque paire se positionne sur le
plan équatorial indépendamment des autres paires. Il se forme à chaque pôle de la cellule un assortiment aléatoire
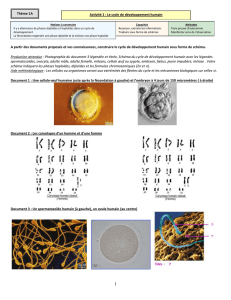
de chromosomes maternels et paternels. Vu qu’il y a 23 paires de chromosomes chez la femme et l’homme, le
brassage interchromosomique permet la formation de 223 gamètes (8,3 millions) avec une répartition différente des
chromosomes maternels et paternels. C'est d'ailleurs au cours de ce brassage que peuvent avoir lieu des erreurs de
répartition des chromosomes (voir plus tard).
La télophase 1 + cytocinèse :
Deux lots haploïdes de chromosomes à deux chromatides sont disposés à chaque pôle de la cellule. Les
nucléoles et les membranes nucléaires se reforment. Les chromosomes se décondensent. La cytocinèse se termine
avec la première division nucléaire et permet la formation de deux cellules filles possédant chacune un lot haploïde
de chromosomes bichromatidiens.
Attention : Il y a plusieurs possibilités de cellules filles suivant que le brassage intrachromosomique (crossing-
over) ait eu lieu et suivant également le brassage interchromosomique. On dit que chaque cellule issue de la méiose
1 et s'apprêtant en entrer en méiose 2 possède une association de chromosomes unique.

b- Méiose 2 ou division équationnelle :
Prophase II Métaphase II Anaphase II Télophase II (+ cytodiérèse)
La deuxième division de la méiose permet la séparation des chromatides sœurs. Elle est semblable à une mitose
avec quelques particularités :
- elle n'est pas précédée d'une réplication de l'ADN
- les chromatides sœurs ne sont pas forcément génétiquement identiques à cause des crossing-over (brassage
intrachromosomique).
Détais de la Méiose 2 :
Prophase II :
Cette phase est identique à la prophase I mais elle est plus brève car les chromosomes sont restés compactés.
phase, une séparation des centrosomes s'effectue. Ainsi, 2 centrosomes génèrent des fuseaux de fibres entre eux et
s'éloignent l'un de l'autre. Les microtubules kinétochoriens s'attachent au kinétochores, à raison d'un kinétochore
par chromatide, de part et d'autre du centromère (de manière à séparer les chromatides).
Métaphase II :
Les chromosomes se placent sur la plaque équatoriale par leur centromère. Leur condensation est maximale.
Anaphase II :
Les chromatides sœurs de chaque chromosome se séparent après rupture au niveau du centromère et migrent
vers les pôles opposés de la cellule tirés par les microtubules. Durant cette phase, les microtubules du fuseau de
division tirent les chromatides sœurs vers les pôles opposées de la cellule.
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%