Lire l`article complet

131
Act. Méd. Int. - Hypertension (10), n° 6, juin 1998
Progrès en hypertension
Mécanismes molé-
culaires de l’activa-
tion des récepteurs
AT1et de l’angio-
tensine II :
conséquences
physiopathologiques
M. Andrejak (Amiens)
D’après la communication de
E. Clauser (Collège de France, Paris)
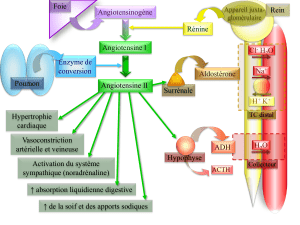
La stimulation des récep-
teurs AT1est responsable
des actions dites classiques
de l’angiotensine II :
contraction du muscle lisse
et sécrétion d’aldostérone.
L’activation d’une voie de
signalisation, mettant en
jeu une protéine Gq, une
phospholipase C, et abou-
tissant, par l’intermédiaire
de l’inositol triphosphate, à
une mobilisation du calcium
intracellulaire et une activa-
tion de protéines kinases C a
été décrite lors de la stimula-
tion des AT1récepteurs.
Les récepteurs AT1(subdivisés en deux
sous-types, AT1A et AT1B, chez le ron-
geur alors qu’un seul récepteur AT1est
connu chez l’homme) ont été clonés
dans de nombreuses espèces. Ce sont
des récepteurs à sept domaines trans-
membranaires (segments hydrophobes
en hélices α reliés entre eux par des
boucles extra- ou intracellulaires)
(figure 1).
Les acides aminés impliqués dans la
fixation de l’angiotensine sont extra-
cellulaires alors que les sites de liaison
pour les anti-AT1impliquent des
acides aminés des domaines trans-
membranaires. Les deux sites de liai-
son sont donc différents sur le plan
moléculaire.
Des travaux de mutagenèse dirigés
pour mieux préciser le couplage
récepteur AT1-protéine G
Pour préciser les séquences d’acides
aminés impliqués dans le couplage aux
protéines G du récepteur AT1, les tra-
vaux se sont appuyés sur les connais-
sances acquises avec les récepteurs adré-
nergiques. Au niveau de ces récepteurs,
un segment distal de la troisième boucle
intracellulaire intervient dans le coupla-
ge récepteur-protéine G.
Sept acides aminés du segment ana-
logue du récepteur AT1ont été rempla-
cés par :
- la séquence correspondante du récep-
Figure 1 : Séquences fonctionnelles du récepteur AT1de l’angiotensine II.
MEP juin 2 15/04/04 10:34 Page 131

Progrès en hypertension
Act. Méd. Int. - Hypertension (10), n° 6, juin 1998
132
teur α1-adrénergique, qui est couplé,
comme le récepteur AT1, à une phos-
pholipase C ;
- la séquence correspondante du récep-
teur ß2-adrénergique, couplé positive-
ment à l’adénylate cyclase ;
- pour finir, la même séquence du
récepteur AT2couplé à des protéines Gi
et à des tyrosine-phosphatases.
Ces récepteurs ont été exprimés dans une
cellule hétérologue. La signalisation y est
modifiée par ces transformations des
récepteurs AT1. Ainsi, l’inositol phospha-
te est induit pour des doses croissantes
d’angiotensine II de façon marquée par
les récepteurs sauvages AT1comme pour
les récepteurs substitués par la séquence
α1-adrénergique, alors que la réponse est
nulle pour les récepteurs chimères AT2et
faible pour les ß2. En ce qui concerne la
production d’AMPc, elle n’est augmentée
sous angiotensine II que pour les récep-
teurs substitués par la séquence ß2. Ces
modifications de couplage et des voies de
signalisation de ces récepteurs chimères
ont des conséquences sur les effets phy-
siologiques de l’angiotensine II. Ainsi,
l’effet mitogène mesuré par l’incor-
poration de thymidine tritiée dans l’ADN
est induit par l’angiotensine II dans les
cellules exprimant le récepteur sauvage
ainsi que dans les cellules chimères expri-
mant le récepteur α1-adrénergique mais
pas dans les cellules contenant les deux
autres types de récepteurs (C-ß2 et C-
AT2) (figure 2).
Donc, ce segment de la troisième boucle
intracellulaire est clairement impliqué
dans le couplage de la protéine G et sur-
tout dans la spécificité de ce couplage (2).
Mécanismes d’activation
moléculaire du récepteur AT1
(ou comment le récepteur passe-t-il
d’un état inactif à un état actif ?)
L’activation passerait par une transcon-
formation des sept segments transmem-
branaires, avec un état inactif du récep-
teur qui aurait une meilleure affinité pour
les agonistes inversés et un état activé qui
aurait une meilleure affinité pour les ago-
nistes, les antagonistes ayant une affinité
identique pour les deux états. Ces chan-
gements de disposition seraient détermi-
nants pour l’activation du récepteur. Des
mutations peuvent bloquer soit dans un
état inactif, soit dans un état activé (acti-
vation dite constitutive) les récepteurs.
Des acides aminés polaires ont été identi-
fiés dans les segments transmembra-
naires dont la mutation inactive le récep-
teur sans modifier la liaison pour les
agonistes peptidiques ni les séquences
intracellulaires. Ces mutations, néan-
moins, empêchent la transformation du
récepteur d’un état inactif vers un état
actif. De même, ont été recherchées des
mutations qui pourraient bloquer les
récepteurs AT1dans un état actif (activa-
tion constitutive à l’instar de ce qui a
d’abord été décrit pour les récepteurs
adré-nergiques). Aucune activation
constitutive n’a été constatée pour les
récepteurs AT1chi-
mères de la troisième
boucle décrits précé-
demment, alors qu’elle
existe pour des muta-
tions de récepteurs α1-
adrénergiques.
Deux groupes seule-
ment (Groblewski et
al., 1997 ; Balmforth
et al., 1997) ont iden-
tifié une activation
constitutive de récep-
teurs AT 1 par muta-
tion au niveau des
domaines transmem-
branaires (1, 4). Il
s’agit des asparagines
111 et 295, qui se
trouvent dans les troi-
sièmes et septièmes
domaines transmem-
branaires. Ces deux acides aminés
auraient des interactions faibles (type
liaisons hydrogène) qui seraient
capables de stabiliser le récepteur dans
un état activé. La mutation de ces acides
aminés activerait constitutivement le
récepteur. Ces mutants ont une affinité
tout à fait normale pour l’angiotensine II.
Par contre, leur affinité pour les antago-
nistes AT1sélectifs est réduite. En ce qui
concerne la signalisation appréciée par
la production d’inositol phosphate, le
niveau de base est augmenté pour les
formes mutantes constitutivement acti-
vées du récepteur AT1, l’angiotensine II
restant cependant toujours capable de
majorer la signalisation. Ces récepteurs
ne seraient donc que partiellement actifs
constitutionnellement.
Existe-t-il des pathologies liées
à une activation constitutive
des récepteurs AT1?
Dans certaines maladies endocriniennes,
Figure 2 : Stimulation par l’angiotensine II de la synthèse
d’ADN.
MEP juin 2 15/04/04 10:34 Page 132

133
des mutations de certains récepteurs ont
été mises en évidence, responsables de
leur activation constitutive. Des muta-
tions des récepteurs à la LH entraînent un
syndrome de puberté précoce. La muta-
tion du récepteur de la TSH peut être à
l’origine d’un adénome toxique de la thy-
roïde.
La possibilité de mutations somatiques
constitutivement activatrices du récep-
teur AT1a été recherchée dans l’adéno-
me de Conn dans le cadre du réseau
COMETE (animé par P. F. Plouin),
cette tumeur bénigne hypersécrétant de
l’aldostérone de façon non régulable.
Par analogie, ce modèle peut être consi-
déré comme un équivalent surrénalien
de l’adénome toxique de la thyroïde.
L’angiotensine II étant l’un des facteurs
majeurs de stimulation de la sécrétion
d’aldostérone, il était logique de sus-
pecter une activation constitutive de ce
récepteur responsable d’une hypersé-
crétion d’aldostérone non régulable.
Cependant, le séquençage du gène de
ce récepteur dans 17 tumeurs de Conn a
permis d’exclure ce mécanisme et la
responsabilité du récepteur AT1(3).
Mécanismes des effets de la stimula-
tion AT1 sur la croissance cellulaire
Si les actions hypertrophiques (aug-
mentation de taille des cellules) de l’an-
giotensine II sur ses différents tissus
cibles sont clairement établies, ses
actions hyperplasiques (augmentation
du nombre des cellules) sont beaucoup
plus débattues et pourraient dépendre
du type cellulaire et de la coopération
d’autres facteurs de croissance.
L’hyperplasie cellulaire pourrait relever
de mécanismes indirects, l’angiotensine
II stimulant au niveau des cellules la
production de facteurs de croissance
qui, de manière autocrine ou paracrine,
favoriseraient la division cellulaire.
L’angiotensine II pourrait aussi agir
directement par ses voies de signa-
lisation. Dans le modèle développé par
l’équipe d’E. Clauser de fibroblastes
surexprimant le récepteur AT1, l’effet de
l’AT1sur la division cellulaire est étudié
par l’incorporation de thymidine tritiée
dans l’ADN (figure 3). Cet effet est très
marqué, dose-dépendant, (alors que
l’angiotensine II est sans effet dans une
lignée CHO non transfectée) et est blo-
qué par un antagoniste AT1spécifique.
Dans ce modèle cellulaire, l’action mito-
gène de l’angiotensine II semble être
médiée par l’activation des protéines
kinases C et la mobilisation du calcium.
L’hypothèse d’un effet indirect par les
facteurs de croissance est peu probable
dans ce modèle. En effet, les deux fac-
teurs de croissance principaux, le FGF et
l’IGF1, qui stimulent très fortement la
division cellulaire, voient leur effet tota-
lement bloqué par des anticorps contre
ces deux facteurs de croissance alors que
ces anticorps n’ont aucune action sur
l’effet mitogène de l’angiotensine II.
La figure 4 montre l’ensemble des
voies potentiellement mises en jeu par
la stimulation AT1. A côté de la phos-
pholipase C, il faut noter qu’il peut y
avoir activation ou inhibition de l’adé-
nylcyclase ou enfin mise en jeu de la
phospholipase A2, ces différentes voies
pouvant intervenir pour moduler la
division cellulaire.
Plus récemment, on a montré que le
récepteur AT1pouvait activer d’autres
voies de signalisation (voie ras des MAP
kinases, voie Jak-STAT des cytokines,
phosphorylation sur la tyrosine de pro-
téines intracellulaires…).
En fait, la voie de signalisation qui
Figure 3 : Récepteur AT1A et croissance cellulaire induite par l’angiotensine II.
MEP juin 2 15/04/04 10:34 Page 133

Progrès en hypertension
Act. Méd. Int. - Hypertension (10), n° 6, juin 1998
134
semble dominer est la voie de la phos-
pholipase C avec ses deux seconds
messagers, diacylglycérol et inositol
triphosphate.
Les conclusions de cette présentation
d’Eric Clauser sont les suivantes :
• Le site de couplage aux protéines G
implique en particulier la troisième
boucle intracellulaire, qui détermine la
spécificité de ce couplage.
• L’activation du récepteur AT1corres-
pond à un changement de conformation
des hélices, qui implique des résidus
polaires des segments transmembra-
naires, dont les mutations entraînent
soit une inactivation, soit une activation
constitutive.
• L’adénome de Conn n’est pas dû à des
mutations somatiques activatrices de la
séquence codante du récepteur AT1.
• L’effet mitogène de l’angiotensine II
met en jeu les récepteurs AT1et
implique, au moins dans certaines cel-
lules, la stimulation directe de la voie
de signalisation de la phospholipase C.
Figure 4 : Signalisation de l’effet mitogène de l’angiotensine II. Références
1) Balmforth A.J., Lee A.J., Warburton P.,
Donnelly D., Ball B. : The conformatio-
nal change responsible for AT1receptor
activation is dependent upon two juxta-
posed asparagine residues on transmem-
brane helices III and VII. J. Biol. Chem.,
1997, 272 : 4245-4251.
2) Conchon S., Barrault M.B., Miserey S.,
Corvol P., Clauser E. : The C-terminal
third intracellular loop of the rat AT1A
angiotensin receptor plays a key role in
G protein coupling specificity and trans-
duction of the mitogenic signal. J. Biol.
Chem., 1997, 272 : 25566-25572.
3) Davies E., Bonnardeaux A., Plouin
P.F., Corvol P., Clauser E. : Somatic
mutations of the angiotensin II (AT1)
receptor gene are not present in aldoste-
rone-producing adenoma. J. Clin.
Endocrinol. Metab., 1997, 82 : 611-615.
4) Groblewski T., Maigret B., Larguier R.,
Lombard C., Bonnafous J.C., Marie J. :
Mutation of Asn 111 in the third trans-
membrane domain of the AT 1A angioten-
sin II receptor induces its constitutive
activation. J. Biol. Chem., 1997, 272 :
1822-1826.
MEP juin 2 15/04/04 10:34 Page 134
1
/
4
100%