STABILITE ET VARIATION DES GENOMES ET EVOLUTION

CH3 : STABILITE ET VARIATION DES GENOMES ET EVOLUTION
INTRODUCTION :
Caractères biologiques, ancestraux ou dérivés, sont déterminés par l’ADN : les séquences protéiques, traduction des
de séquences de nucléotides, permettent l’activité cellulaire et la mise en place des organes. Les innovations
biologiques, concernant les organes, les cellules ou les molécules, dépendent des innovations génétiques.
Comment apparaissent de nouveaux allèles et de nouveaux gènes ? Comment les caryotypes peuvent-ils
être à la fois stables dans l’espèce et variables au cours du temps ? Comment de nouveaux génotypes
peuvent-ils se conserver dans une population ?
PLAN
I. LES INNOVATIONS GENETIQUES
A Nouveaux allèles, B Nouveaux gènes, C Instabilité de l’ADN, D Phénotype modifié
II. STABILITE DU CARYOTYPE DANS UNE ESPECE
A Alternance de phases, B Méiose, C Anomalies chromosomiques
III. BRASSAGE GENETIQUE LORS DE LA REPRODUCTION SEXUEE
A Conséquence diploïdie, B Recombinaison méiose, C Amplification à la fécondation
IV. INNOVATIONS GENETIQUES ET EVOLUTION DES ESPECES
A Avantage sélectif, B Polymorphisme équilibré, C Mutations neutres, D Mutation développement
CONCLUSION

I : LES INNOVATIONS GENETIQUES retour plan
On sait que les individus d’une espèce ou d’une population ont le même caryotype et le même génome. On peut
penser qu’une population possède un caryotype et un génome stable.
Comment des innovations génétiques peuvent-elles apparaître si l’espèce est stable ? Qu’est-ce qu’une
innovation génétique ? Quelle explication l’étude du génome peut-elle apporter ?
A. APPARITION DE NOUVEAUX ALLELES DANS UNE POPULATION
1. Polyallélisme à l’origine de la diversité des individus
Dans une même population, les gènes existent généralement sous nombreux allèles : ils sont poly-alléliques.
Ex : gène CFTR (mucoviscidose) : 100 allèles ; gène HLA : 108 allèles ; 2 gènes sur 3 sont poly-alléliques.
Pour chacun de ces gènes, chaque individu possède 2 allèles, parmi les nombreux possibles. Les couples d’allèles
sont très variables : un individu est génétiquement unique. Les individus d’une même population ont même
génome mais diffèrent par leurs couples d’allèles.
2. Polymorphisme génique
Si, au moins 2 allèles ont une fréquence supérieure à 1%, on dit que ces gènes poly-alléliques sont polymorphes,
ou possèdent un polymorphisme génique, ce qui signifie qu’ils possèdent plusieurs formes alléliques non rares.
Ex : ABO : chaque forme à une forte fréquence : A 30%, B 10%, O 60% : le gène ABO est polymorphe
3. Les mutations à l’origine du polyallélisme
Les mutations ponctuelles, de 1 ou quelques nucléotides, modifient la séquence de nucléotides par, addition,
délétion (= mutations décalantes), ou par substitution : elles sont à l’origine de nouveaux allèles. Au sein d’une
espèce, le polymorphisme de l'ADN résulte de l’accumulation de mutations au cours des générations.

B. APPARITION DE NOUVEAUX GENES DANS UNE ESPECE retour plan
1. Les gènes homologues forment des familles multigéniques
Certains gènes possèdent des séquences ressemblantes. On pense qu’une similitude de 20 % entre des séquences
n’est pas due au hasard et signifie qu’elles dérivent d’un même gène ancestral. Ces gènes dits homologues
forment une famille multigénique :
Ex : Famille des globines : http://www.inrp.fr/Acces/biotic/evolut/mecanismes/globines/html/synthese.htm#1
2. Duplications et mutations à l’origine des gènes homologues
Les gènes homologues viennent de la duplication d’un gène ancestral, suivie de mutations aléatoires.
L’accumulation des mutations provoque des dérives génétiques indépendantes et les 2 exemplaires deviennent des
gènes distincts. Plus des gènes sont âgés, plus les mutations sont accumulées, plus la divergence génétique est
forte. La connaissance du rythme de mutation d’une molécule permet d’évaluer l’âge de duplication du gène
ancestral.
Les duplications sont des mutations étendues et résultent du doublement d’un fragment de chromosome suivi
d’une transposition (= déplacement sur le même chromosome ou sur un autre chromosome).
Attention : Duplication = doublement d’un gène ;
réplication de l’ADN, doublement d’un chromatide.
C. L’INSTABILITE DE L’ADN A L’ORIGINE DES MUTATIONS
1. Origine des mutations ponctuelles = elles viennent d’erreurs d’appariement spontanées (= indépendantes
du milieu), aléatoires (= au hasard) et rares, lors de la réplication.
Les substances mutagènes (dioxine), les rayonnements X, UV favorisent ces mutations ponctuelles.
2. Origine des duplications : elles viennent d’échanges inégaux entre des chromosomes.

D. LES MUTATIONS PEUVENT MODIFIER LE PHENOTYPE ET SE TRANSMETTRE retour plan
Les mutations ont des conséquences variables suivant leur place dans la séquence, leur signification dans le code
génétique et la cellule qui les porte :
L’activité de la protéine est modifiée :
si la mutation est « faux sens » : elle provoque un changement d’acide aminé dans le site actif
si la mutation est « non sens » : elle créé un codon stop qui raccourcit la protéine.
Dans les gènes homéotiques, elles peuvent entraîner de grands changements. Ex : Gène UBX de drosophile.
Le site actif ne change pas d’activité :
si la mutation est « neutre » : la mutation « faux sens » est hors du site actif.
si la mutation est « silencieuse » : le codon muté signifie le même acide aminé dans le code génétique.
Elles complexifient le génome : Les duplications suivies de mutations, augmentent la quantité d’information
génétique disponible : la quantité d’information génétique augmente. Exemple des globines ↑
Exemple des opsines : http://www.inrp.fr/Acces/biotic/evolut/mecanismes/opsines/html/resultatscomparaison.htm
Elles se transmettent si elles sont dans les cellules germinales (= C/ sexuelles), et participent à la fécondation.
Les mutations somatiques ne concernent que les organes non reproducteurs et restent dans l’individu.
BILAN
Les mutations ponctuelles sont à l’origine de la polyallélie ; les duplications, mutations étendues, à l’origine
des familles multigéniques. Ces mutations apportent la diversification génétique individuelle et la
complexification du génome de l’espèce, à condition qu’elles soient transmises par les cellules germinales et
si elles ne sont pas trop désavantageuses pour l’individu. Les mutations sont aléatoires et leur nature ne
dépend pas de l’environnement.

II : STABILITE DU CARYOTYPE DE L’ESPECE retour plan
Lors de la fécondation, les ovules et les spermatozoïdes fusionnent : leur matériel génétique s’unit.
Comment la stabilité du caryotype est-elle maintenue dans l’espèce au cours de la reproduction ?
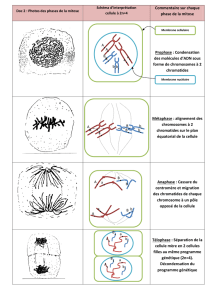
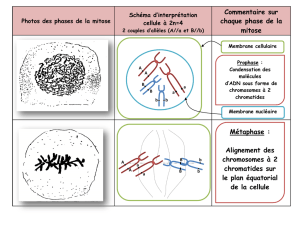
A. ALTERNANCE DE PHASES DANS LE CYCLE DE REPRODUCTION
1. Cycle de reproduction d’un mammifère = cycle diploïde
Toutes cellules de l’organisme dérivent d’un œuf diploïde (2N chromosomes). (Œuf = zygote)
Dans les gonades, certaines cellules diploïdes deviennent des gamètes haploïdes (N chromosomes), juste avant la
fécondation. La phase diploïde est dominante : 2N de l’œuf à l’adulte ; N aux gamètes
Individu femelle adulte
Individu mâle adulte 2N
MEIOSE RC
PH haploïde = gamètes N
Gamètes mâles Gamète femelle
FECONDATION
Œuf 2N
 6
7
8
9
10
11
12
13
14
15
6
7
8
9
10
11
12
13
14
15
1
/
15
100%