ecole nationale veterinaire d`alfort

ECOLE NATIONALE VETERINAIRE D’ALFORT
Année 2002
CONTRIBUTION A LA MISE AU POINT D’UNE
ELECTRODE DTL POUR LE RECUEIL DE
L’ELECTRORETINOGRAMME CHEZ LE CHIEN
THESE
pour le
DOCTORAT VETERINAIRE
Présentée et soutenue publiquement
Devant
LA FACULTE DE MEDECINE DE CRETEIL
le
par
Arnaud LELIEVRE DU BROEUILLE
Né le 24 octobre 1976 à Boulogne sur mer (Pas de Calais)
JURY
Président :M
Professeur à la Faculté de Médecine de Créteil
Membres
Directeur : M CLERC
Professeur à l’Ecole Nationale Vétérinaire d’Alfort
Assesseur :M MICHAUX
Professeur à l’Ecole Nationale Vétérinaire d’Alfort
0

A notre jury de thèse,
Monsieur le Professeur,
Professeur à la faculté de médecine de Créteil,
Qui nous a fait l’honneur de présider notre jury de thèse,
Hommage respectueux.
Monsieur le Professeur Clerc,
Professeur à l’Ecole Nationale Vétérinaire d’Alfort,
Pour nous avoir suggéré ce travail
Qu’il trouve ici l’expression de notre profonde reconnaissance.
Monsieur le Professeur Michaux
Professeur à l’Ecole Nationale Vétérinaire d’Alfort,
Qui a accepté de faire partie de notre jury de thèse et de juger ce travail,
Qu’il accepte nos sincères remerciements.
1

Au Docteur Sabine Chahory,
Qui nous a aidée à la réalisation des examens électrorétinographiques,
Qu’elle trouve ici l’expression de nos sincères remerciements.
Aux docteurs Trio et Vanore,
Qui nous ont apportées toutes leurs aides techniques,
Qu’elles trouvent ici l’expression de toute notre gratitude.
Aux personnels du Museum d’Histoire naturelle,
Qui ont accepté que nous examinions ses microcèbes,
Qu’ils reçoivent ici l’expression de toute notre gratitude.
2

A mes parents,
Pour leur amour et leur soutien constant,
Qui m’ont toujours encouragé dans cette voie,
Qu’ils soient remerciés pour tout ce qu’ils ont fait pour moi.
A ma sœur Laëtitia et à son fiancé Emmanuel,
A toute ma famille,
A mes amis,
3

TABLE DES MATIERES
TABLE DES MATIERES………………………………..………………………..……..…1
INTRODUCTION…………………………………………...………………..……………..5
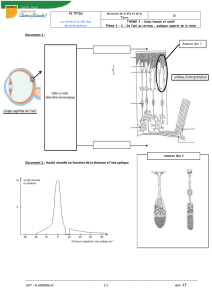
ANATOMIE-HISTOLOGIE-PHYSIOLOGIE DE LA RETINE…..….....…..………….7
A. Anatomie et histologie de la rétine...…………...……………………..…………8
1. Développement……………...…………………………………..………….8
2. Histologie de la
rétine…………………………………………..…………..8
2.1. L’épithélium
pigmentaire…………………………...…………….8
2.2. Les photorécepteurs…………………………………...………….9
2.3. La membrane limitante
externe……………………..….…………9
2.4. La couche nucléaire
externe…………………………...………….9
2.5. La couche plexiforme
externe………………………...…………..9
2.6. La couche nucléaire
interne………………………………..…….10
2.7. La couche plexiforme
interne……………………………..……..10
2.8. La couche des cellules
ganglionnaires…………………..……….10
2.9. La couche des fibres
nerveuses…………………………..………10
2.10. La membrane limitante
interne……………………..…………..10
B. Etude spécifique des photorécepteurs….……...…………………..…………...12
1. L’expansion externe………...…………………….…………..………...…12
1.1. Le segment externe…………………………………..………….12
1.2. La zone
intermédiaire………………………………...………….12
1.3. Le segment interne……………………………..………………..12
2. L’expansion interne…………………………………………………..…...13
2.1. Le corps ou couche nucléaire
externe………………………..….13
2.2. La terminaison synaptique ou
pied…………………………..….13
C. Physiologie rétinienne………………..……...…………………………….….....14
1. Les pigments rétiniens………...…………………………………………..14
1.1. La rhodopsine…………………………………………...……….15
1.2. Les iodopsines………………...…………………………………15
2. La transduction
visuelle..……………………………………………..…...15
4
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
1
/
84
100%