Revue Leptine et contrôle hypothalamique de la reproduction

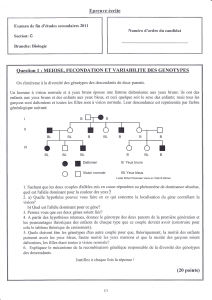
Revue
Leptine et contrôle hypothalamique
de la reproduction
Leptin and hypothalamic control of reproduction
Sébastien G. Bouret
1,2
Émilie Caron
1
Sophie Steculorum
1
Yuko Ishii
1,2
Christelle Sachot
1
1
Inserm, U837, Centre de recherche
Jean-Pierre Aubert, Développement et
plasticité du cerveau postnatal,
Université de Lille 2, 1 place de Verdun,
59045 Lille Cedex, France
2
The Saban Research Institute,
Neuroscience Program, Childrens Hospital
of Los Angeles, University of Southern
California, 4650 Sunset Boulevard,
MS #135, Los Angeles, CA 90027, USA
Résumé.La fertilité est étroitement régulée par la nutrition et la disponibilité des réserves
énergétiques. Ainsi, les hormones adipocytaires comme la leptine tiennent une place centrale
dans la mise en place et la régulation de la fonction de reproduction. Outre son action directe
au niveau des cellules gonadotropes de l’hypophyse et des gonades, la leptine agit également
sur le cerveau neuroendocrine par l’intermédiaire de ses récepteurs localisés au niveau de
l’hypothalamus. Cette revue générale met l’accent sur les mécanismes hypothalamiques de
contrôle de la reproduction par la leptine et aborde certaines pathologies où une dysrégula-
tion de la leptinémie est associée à des troubles de la reproduction.
Mots clés : hormones, nutrition, GnRH, développement
Abstract.Fertility is gated by nutrition and the availability of stored energy reserves. Thus, the
adipocyte-derived hormone leptin appears to play a pivotal role in the onset and regulation of
reproductive function. In addition to its direct action at the pituitary and gonad levels, recent
data has defined a core circuitry in the hypothalamus that appears to mediate some of the
effects of leptin on reproductive function. This review focuses on the role of leptin in the
hypothalamic control of reproduction and discusses about reproductive pathologies associa-
ted with altered leptin levels.
Key words: hormones, nutrition, GnRH, development
L
es impératifs biologiques primor-
diaux imposés à un organisme sont
d’abord d’assurer sa survie et ensuite de
pérenniser son espèce. Ainsi, le système
reproducteur des mammifères est extrê-
mement sensible à la disponibilité des
ressources énergétiques dans l’environ-
nement. Des modifications du statut
énergétique, comme la restriction ali-
mentaire ou l’obésité, peuvent induire
des remaniements de l’axe hypotha-
lamo-hypophysio-gonadique (HPG) et
conduire, par exemple, à l’arrêt de la
sécrétion de l’hormone lutéinisante (LH)
ou altérer la mise en place de la puberté.
Les intermédiaires métaboliques mis en
jeu dans les interactions nutrition/
reproduction restent encore mal connus
mais les travaux réalisés depuis ces dix
dernières années suggèrent que l’hor-
mone adipocytaire leptine pourrait être
l’un des facteurs capable de réaliser le
lien entre l’équilibre énergétique et la
reproduction en agissant, au moins en
partie, au niveau de l’hypothalamus
neuroendocrine.
La stabilité du poids corporel et de
la masse adipeuse doit s’obtenir par
l’équilibre des dépenses (besoins phy-
siologiques) et des apports énergéti-
ques (alimentaires). Basé sur cette ob-
servation, Kennedy a imaginé un
mécanisme permettant le maintien du
poids corporel à un niveau constant
selon lequel un signal émanant des
stocks énergétiques stimulerait des
changements compensatoires de la
prise alimentaire et des dépenses éner-
gétiques afin que la masse adipeuse
reste à un niveau stable [1, 2]
.
Ce point
de vue a été confirmé par des études
ultérieures montrant que le gain de
mt Médecine de la Reproduction, Gynécologie Endocrinologie 2008 ; 10 (2) : 74-84
médecine thérapeutique
Médecine
de la Reproduction
Gynécologie
Endocrinologie
Tirés à part : S.G. Bouret
doi: 10.1684/mte.2008.0147
mt Médecine de la Reproduction, Gynécologie Endocrinologie, vol. 10, n° 2, mars-avril 2008
74
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

poids obtenu par gavage avait pour conséquence une dimi-
nution volontaire de la prise alimentaire, une augmentation
des dépenses énergétiques et donc un éventuel retour au
poids initial. À l’inverse, la mise à jeun ou la lipectomie
stimulent la prise alimentaire et diminuent les dépenses
énergétiques afin de restaurer le poids et la masse adipeuse
initiaux [3, 4]
. Cependant, les signaux véhiculant la possi-
ble interaction entre le tissu adipeux et le comportement
alimentaire sont restés inconnus pendant plusieurs an-
nées. Les études de Hervey ont fourni d’importantes don-
nées sur la nature de ces signaux potentiels [5, 6]. La mise
en parabiose (circulation croisée) d’un rat dont la partie
médiobasale de l’hypothalamus a été détruite avec un rat
non lésé provoque la perte d’appétit et de poids du rat
intact. En revanche, les rats lésés continuent à gagner du
poids même s’ils sont mis en parabiose avec un rat normal
ou un autre rat présentant les mêmes lésions. Ces études
suggèrent que les rats obèses dont la partie médiobasale
de l’hypothalamus a été détruite produisent un facteur de
« satiété » circulant provoquant la perte d’appétit chez les
rats parabiotiques non lésés. Par ailleurs, l’absence de
réponse des rats lésés est en accord avec l’existence d’un
centre de la satiété comme proposé lors d’études antérieu-
res [7]. Des travaux ultérieurs réalisés chez la souris
« obèse » (ob) ont contribué à l’évolution des connaissan-
ces sur ce facteur impliqué dans la satiété. La mise en
parabiose d’une souris sauvage avec une souris « obèse »
(ob) permet aux souris obèses de perdre du poids, alors
que la parabiose entre une souris sauvage et une autre
souche de souris obèse, la souris « diabétique » (db),
provoque une hypophagie et une perte de poids chez la
souris sauvage [8, 9]. Ces travaux mettent en lumière
l’implication du locus ob dans la production d’un facteur
de satiété, alors que le locus db code pour un composant
requis dans le mécanisme de réponse au facteur de satiété.
Ces prédictions basées sur des expériences de parabiose
ont été confirmées par le clonage des gènes ob et db au
milieu des années 1990. Le gène murin ob (leptine),
découvert grâce au clonage positionnel, code pour un
ARNm de 4,5 kb contenant un cadre ouvert de lecture
hautement conservé de 167 acides aminés [10]. Le produit
du gène ob est synthétisé principalement par le tissu
adipeux blanc, en quantité proportionnelle à la masse
adipeuse, et il est ensuite libéré dans le sang [11]. Rapide-
ment après le gène de la leptine, le gène db codant pour
son récepteur de forme longue a été cloné [12, 13]. Ce
récepteur, appelé LRb (ou encore Ob-Rb), est un récepteur
transmembranaire qui présente une forte homologie avec
la famille des récepteurs de cytokines de classe I [14-16].
Rôles de la leptine
sur la fonction de reproduction
En même temps qu’était décrite l’obésité morbide des
souris ob/ob, Ingalls et al. faisaient état de la stérilité de
celles-ci [17]. De façon intéressante, les femelles déficien-
tes en leptine restent stériles quel que soit leur poids [18].
De plus, malgré un développement sexuel précoce nor-
mal, les femelles ob/ob et db/db restent indéfiniment
prépubères sans jamais présenter d’ovulation ni de cycle
estral. Témoins du mauvais fonctionnement de leur axe
gonadotrope, leurs organes sexuels ne se développent pas
et leurs taux d’hormones stéroïdiennes sont anormale-
ment bas [19]. De façon remarquable, l’administration de
leptine restaure les déficits de la fonction de reproduction
observés chez les souris ob/ob [20]. L’ensemble de ces
travaux démontrent que des déficiences en leptine ou en
son récepteur induisent des troubles de la reproduction et
ont emmené de nombreuses équipes à s’intéresser au rôle
de cette hormone dans la survenue de la puberté et dans le
contrôle de l’ovulation.
Leptine et puberté
La puberté chez les mammifères est physiologique-
ment liée aux ressources énergétiques de l’individu. En
accord avec cette observation, les travaux de Kennedy et
Mitra ainsi que les études de Frisch et al. ont mis en
évidence que la survenue de la maturation sexuelle est
associée au poids corporel et au contenu du tissu adipeux
[21, 22]. Plus précisément, le régime alimentaire ou l’acti-
vité physique exercent une profonde influence sur la
maturation sexuelle de nombreuses espèces incluant le
rat, le mouton et l’homme [23-25]. Il est important de
souligner que les niveaux de leptine plasmatique sont
étroitement liés aux apports alimentaires et à l’activité
physique. Chez la souris, un pic de leptine postnatale a
lieu entre le 8
e
et le 12
e
jour de vie postnatale [26]. Ces
niveaux élevés de leptine précèdent le pic de FSH se
produisant au 12
e
jour postnatal (P12), au cours de la
phase infantile de la puberté, et le début de la sécrétion
d’estradiol à P14 [26, 27]. Chez l’humain, la survenue de
la puberté est également précédée par une augmentation
du taux de leptine circulante, ce qui pourrait constituer un
signal potentiel du déclenchement de la puberté [28].
Chez les garçons, les taux de leptine atteignent leur apex
lors de la période précédant la survenue de la puberté
(stades Tanner précoces), puis chutent progressivement à
mesure que la testostéronémie augmente pour retrouver
un niveau de base [29-33]. Les filles, quant à elles, béné-
ficient d’une augmentation constante des taux de leptine
circulante au cours du processus pubertaire [29, 31, 30,
33]. Ceci se fait en concordance avec une augmentation
des taux d’œstrogènes circulants [34]. Il est intéressant de
noter qu’il existe chez les filles une corrélation entre la
mise en place de la ménarche et les taux de leptine
sérique : une enfant d’âge pubertaire présentant un taux
de leptine circulante d’1 ng/mL supérieur à celui d’une
autre enfant de la même tranche d’âge verra la survenue
mt Médecine de la Reproduction, Gynécologie Endocrinologie vol. 10, n° 2, mars-avril 2008 75
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

de la ménarche apparaître un mois plus tôt que cette
dernière [35]. Ceci suggère qu’une concentration seuil de
leptine est nécessaire pour la survenue de la ménarche
chez les filles. De nombreuses études ont ainsi été réali-
sées, notamment chez le rongeur, afin de déterminer si la
leptine était capable d’induire à elle seule la survenue de
la puberté chez les animaux prépubères. Cependant, les
résultats de ces travaux n’ont pu apporter d’éléments
conclusifs. En effet, tandis que certaines équipes rappor-
tent que l’administration quotidienne de leptine dès le
sevrage accélère la maturation sexuelle et la survenue de
la puberté chez les souris femelles sauvages [20, 36, 37],
d’autres indiquent que des injections de leptine chez
l’animal prépubère n’influencent pas la survenue de la
puberté [38, 39]. Une importante question méthodologi-
que sépare cependant les deux études : seules les équipes
qui ont utilisé des doses de leptine ne provoquant pas de
perte de poids ont observé une avancée de la puberté,
confirmant que le poids serait un facteur déterminant dans
la survenue de la puberté [21, 22]. Ainsi, la leptine semble
jouer un rôle essentiel dans le déclenchement de la pu-
berté, mais il s’agirait seulement d’un rôle permissif puis-
que ce n’est qu’une fois un seuil de concentration dépassé
que la puberté peut avoir lieu.
Leptine et fertilité chez l’adulte
Les premières études réalisées chez des souris adultes
montrent que l’injection périphérique de leptine chez des
souris à jeun pendant 48 heures permet aux femelles de
maintenir un cycle régulier allant jusqu’à l’ovulation et
atténue, chez le mâle, la chute de testostérone induite par
la privation alimentaire [40]. Des études complémentaires
ont permis de préciser les mécanismes permettant à la
leptine d’agir sur l’axe gonadotrope (figure 1). Chez des
rattes ovariectomisées, supplémentées ou non en estrogè-
nes et mises à jeun pendant 48 heures, l’injection de
leptine permet de maintenir leur pulsatilité LH normale-
ment supprimée par le jeûne [41, 42]. L’ensemble de ces
travaux soulignent l’importance de la leptine dans le fonc-
tionnement de l’axe gonadotrope. Cependant, il restait à
déterminer à quel niveau de cet axe la leptine exerçait son
action.
Rôle du cerveau dans la médiation
des effets de la leptine
sur la reproduction
En plus de son action directe au niveau des cellules
gonadotropes de l’hypophyse et des gonades (actions non
détaillées dans le cadre de cette revue) [43-45], la leptine
est capable d’agir au niveau du cerveau pour moduler
l’activité de l’axe gonadotrope. Des injections de leptine
dans le troisième ventricule cérébral chez des souris ova-
riectomisées et supplémentées en œstrogènes induisent
une augmentation des taux de LH et de FSH plasmatiques
[46]. De plus, l’injection centrale de leptine
(116-130)
, qui
est un fragment actif de la molécule native, chez des rats
adultes mâles à jeun permet d’augmenter la fréquence et
l’amplitude des pulses de LH ainsi que la sécrétion
moyenne et nette de LH [47]. L’ensemble de ces données
confirment l’hypothèse que la leptine pourrait stimuler
l’axe gonadotrope via une action sur le système nerveux
central. Il semblerait notamment que les effets de la leptine
sur la fonction de reproduction impliquent l’hypothala-
mus et soient liés à des modifications de la sécrétion de
GnRH.
Leptine et sécrétion de GnRH
Plusieurs travaux suggèrent que la leptine exerce une
action stimulatrice sur la sécrétion de GnRH hypothalami-
que. Des expériences réalisées in vitro montrent en effet
qu’à des concentrations subnanomolaires, la leptine sti-
mule la libération de GnRH à partir d’explants incluant le
noyau arqué et l’éminence médiane de rat [46]. Cepen-
dant, en concentration micromolaire, la leptine perd son
effet positif sur la libération de GnRH et, au contraire,
induit une diminution de la production de GnRH par
rapport aux contrôles. Ces résultats ont été confirmés par
des expériences réalisées sur des lignées cellulaires de
neurones immortalisés sécrétant la GnRH (GT1-7), qui
expriment le récepteur à la leptine. Sur cette lignée cellu-
laire, la leptine stimule la libération de GnRH quand elle
est en faible concentration (10
-12
à10
-10
M) mais pas en
forte concentration (10
-8
M) [48]. Cependant, la question
de savoir si la leptine agit directement sur les neurones à
GnRH reste entière dans la mesure où les études de double
marquage menées chez le rongeur et le primate n’ont pas
réussi à mettre en évidence une expression de récepteur à
la leptine par les neurones à GnRH in vivo [49] (figure 2).
L’hypothèse de réseaux neuronaux servant d’intermédiai-
res entre la leptine et les neurones à GnRH reste donc la
plus probable.
Caractérisation des cibles
hypothalamiques directes
de la leptine
Dans la mesure où la souris présentant un récepteur
LRb non fonctionnel (souris db/db) reproduit quasiment à
l’identique le phénotype de la souris déficiente en leptine
(ob/ob), la forme longue du récepteur à la leptine semble
requise pour véhiculer la majeure partie des effets physio-
logiques et neuroendocriniens de la leptine. De plus, le
cerveau apparaît comme un site d’action privilégié pour
Revue
mt Médecine de la Reproduction, Gynécologie Endocrinologie, vol. 10, n° 2, mars-avril 2008
76
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

véhiculer les effets de la leptine, via LRb, sur la fonction de
reproduction puisque la restauration d’un LRb fonctionnel
spécifiquement dans les neurones par transgenèse permet
de restaurer une fertilité normale chez les animaux db/db
mâles comme femelles [50]. Dans le cerveau, la forme
longue du récepteur à la leptine (LRb) est essentiellement
localisée dans l’hypothalamus avec la plus forte expres-
sion retrouvée dans les parties ventrobasales et ventromé-
diales de cette région du cerveau [51, 52]. Ainsi, l’ARNm
de LRb est localisé dans de nombreux noyaux du cerveau
impliqués à la fois dans la régulation de la prise alimen-
taire et le contrôle de la fonction de reproduction, comme
les noyaux arqués, ventromédians et prémammillaires
ventraux. Les travaux de Keen-Rhinehart et al. ont
confirmé l’importance des neurones du noyau arqué dans
la médiation des effets neuroendocrines de la leptine en
normalisant les cycles œstraux de rats Zucker (souche de
rats présentant une mutation de LRb) suite à la restauration
d’un LRb fonctionnel spécifiquement dans leur noyau
arqué [53]. L’expression de LRb ne se limite pas à la partie
médiobasale de l’hypothalamus mais est également re-
trouvée dans d’autres noyaux hypothalamiques tels que le
noyau dorsomédian et l’aire hypothalamique latérale. La
fonctionnalité de LRb dans l’ensemble de ces noyaux a été
confirmée par la capacité de la leptine à induire la phos-
phorylation de STAT3 (pSTAT3, l’une des voies de signali-
sation activée suite à la fixation de la leptine sur LRb) ou
l’expression du proto-oncogène cFos [54, 55]. Un signal
d’hybridation du LRb plus modéré est également observé
au niveau de la région préoptique de l’hypothalamus, où
Neurones
à GnRH
Région
préoptique
Noyau Arqué Cerveau
Hypothalamus
Hypophyse
GnRH
LH, FSH
Cellules
gonadotropes
Ovaires
Testicules
Adipocytes
Leptine
plasmatique
Gonades
Figure 1. Régulation par la leptine de l’axe hypothalamo-hypophyso-gonadique lorsque les conditions nutritionnelles et métaboliques sont
adéquates. Outre son action directe au niveau des cellules gonadotropes de l’hypophyse et des gonades, la leptine (hormone sécrétée
par le tissu adipeux blanc) peut également agir directement au niveau de l’hypothalamus (plus spécifiquement sur les neurones du noyau
arqué) pour stimuler la sécrétion de GnRH. Cette augmentation des taux de GnRH induite par la leptine peut stimuler la sécrétion
d’hormones lutéinisante (LH) et folliculostimulante (FSH) par l’adénohypophyse et ainsi influencer l’ovulation ou la spermatogenèse.
mt Médecine de la Reproduction, Gynécologie Endocrinologie vol. 10, n° 2, mars-avril 2008 77
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

les corps cellulaires des neurones à GnRH sont localisés. Il
est néanmoins important de souligner que même si le LRb
est majoritairement exprimé dans l’hypothalamus au ni-
veau central, d’autres régions du CNS, en particulier le
thalamus et le tronc cérébral, sont également connues
pour exprimer ce récepteur. L’absence d’expression de
LRb dans les neurones à GnRH [49] soulève l’hypothèse
de l’existence de réseaux neuronaux permettant de véhi-
culer transsynaptiquement l’action de la leptine sur les
neurones à GnRH.
Région préoptique
Noyau arqué/EM
GnRH+RL
3V
3V
NA
em
Figure 2. Distribution des corps cellulaires et terminaisons nerveuses contenant la GnRH (fluorescence rouge), et des neurones
exprimant le récepteur à la leptine (fluorescence verte) dans la région préoptique (haut) et au niveau de la région du noyau arqué et de
l’éminence médiane (bas). À noter l’absence de colocalisation entre les neurones à GnRH et les neurones contenant le récepteur à la
leptine. em, éminence médiane ; NA, noyau arqué ; 3V, troisième ventricule.
Revue
mt Médecine de la Reproduction, Gynécologie Endocrinologie, vol. 10, n° 2, mars-avril 2008
78
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%