Correction LIBAN 2010 1ère partie

Correction LIBAN 2010
1ère partie : immunologie
Intro : séropositivité = présence d’AC dans le sang d’un individu infecté. la spécificité des AC dirigés contre les Ag
viraux (glycoprotéine GP120 par ex) permet détection de l’infection au VIH par le test ELISA
Structure des AC. (Autre nom de les AC : les Igg =Immunoglobulines)
Schéma exigé type :
Une molécule de nature protéique/4 chaînes
polypeptidiques associées par liaisons non
peptidiques/un site de reconnaissance et de
fixation de l’ag avec une configuration spatiale
« unique »/ une zone constante permettant
aux AC de se lier à d’autres agents de
l’immunité.
La structure de l’AC permet sa liaison avec un Ag spécifique. Ainsi la réalisation du test ELISA
détermine spécifique la séropositivité au VIH car ce test est basé sur la formation de
complexes immuns :
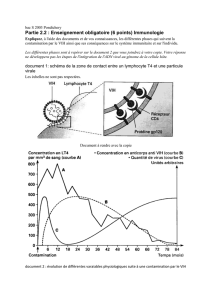
Production des AC en réponse à l’infection au VIH :
Avant l’infection, les AC anti VIH n’existent pas mais des lymphocytes B spécifiques préexistent à l’infection :
l’organisme est capable de produire une infinie diversité de LB différents les 1 des autres par leur « récepteur B »
(autre nom = AC membranaire). Cette diversité est permise par un mécanisme génétique de recombinaison
somatique aléatoire entre les gènes codants pour la synthèse des AC.
L’infection met en circulation dans le sang des particules virales qui peuvent se multiplier en infectant des cellules
immunitaires, les LT4.
La reconnaissance entre le récepteur B d’un lymphocyte B spécifique circulant et une particule virale va enclencher
la réponse immunitaire :
Le LB sélectionné /activé se multiplie (sélection clonale) et produit une grande quantité de LB tous
identiques et tous spécifiques du même Ag
Les LB activés se différencient en cellules sécrétrices d’AC : les plasmocytes. Ces cellules organisées pour
une intense synthèse de protéine (réticulum endoplasmique, app. de golgi) réalisent la sécrétion par
exocytoose d’AC qui vont neutraliser les particules virales circulantes en formant des complexes immuns
spécifiques. Les virus ainsi neutralisé ne peuvent plus contaminer de nouvelles cellules.
Cette phase de production d’AC va durer plusieurs semaines (séroconversion)
L’amplification de cette réponse immunitaire spécifique est contrôlée par des messagers chimiques, les
interleukines produits par les LT4 : c’est une des raisons de l’impossibilité qu’à notre organisme de se
débarrasser du VIH
Conclusion : la réponse immunitaire productrice d’AC spécifique permet de neutraliser spécifiquement les
particules virales (et des les éliminer par la phagocytose). Cependant, cette réponse est insuffisante vis à vis du VIH
car : les AC ne sont pas actifs contre les cellules infectées qui restent des « réservoirs » de VIH et car ce virus mute
fréquemment ce qui rend les AC produits rapidement inutilisables. L’évolution terminale de l’infection au VIH est le
SIDA et le décès.

2ème partie-1 : Datation
NB : La coupe présentée montre différents dépôts sédimentaires et événements tectoniques.
Seuls ceux mentionnées dans la question sont à dater par l’utilisation des principes de la datation relative !
Le dépôt sédimentaire Jurassique s’est mis en place postérieurement aux dépôts du Trias et avant les dépôts du
Crétacé qui l’encadrent : c’est le principe de superposition.
Toutes les failles Fa, Fb et Fc recoupent les dépôts jurassiques : ces failles sont toutes postérieures au jurassique
(c’est le pcp de recoupement).
On constate que la faille Fa recoupe et déplacent la série de failles contemporaines Fb : la faille Fa est donc
postérieure à Fb.
On constate dans la région SSE de la coupe que de part et d’autre de la faille Fc on retrouve la faille Fa « décalée » :
la faille Fc est donc postérieure à Fa.
On peut donc proposer la chronologie suivante pour les 4 événements géologiques demandés :
1. Dépôts sédimentaires du Jurassique
2. Série de failles Fb
3. Faille Fa
4. Faille Fc.

2ème partie-2 : Parenté
Le groupe des Téthythériens (qui comporte les
Proboscidiens actuels et fossiles et le Lamantin) est
délimité en rouge sur le schéma suivant (il exclut les
Suidés).
En bleu, le dernier ancêtre commun de ces mammifères
dont on cherche à déterminer les caractères.
Utilisons les pcp de la phylogénèse pour répondre au
problème posé.
Utilisons le document 1 pour définir les caractères du DAC des Téthythériens (« portrait ») :
On sait que les caractères de l’AC commun d’un groupe sont les caractères dérivés partagés par TOUS les
membres de ce groupe.
- Les états dérivés « présence de poils », « présence de glandes mammaires », « présence d’un amnios »
existent chez le sanglier et les 2 Téthythériens : ces caractères existent chez le DAC des Téthythériens mais
ils ne sont pas exclusifs de ce groupe
- Les états dérivés « perte des canines de la mâchoire inférieure » et « « position reculée des fosses
nasales »existent chez l’éléphant mais pas chez le Lamantin : ces états dérivés n’existent pas chez le DAC
des Téthythériens.
- L’état dérivé « forme en goutte de l’os ectotympanique » existe chez le Lamantin mais pas chez
l’Eléphant : cet état dérivé n’existe pas chez le DAC des Téthythériens.
- L’état dérivé « orbite avancée par rapport à l‘axe A-P du crâne » existe exclusivement chez le lamantin et
l’éléphant et non chez le sanglier : il s’agit d’un caractère apparu chez le DAC des Téthythériens
Bilan : le DAC des Téthythériens possédait des poils, des glandes mammaires (comme tous les mammifères), un
amnios (comme tous les Amniotes) et des orbites avancées.
Utilisons le document 2 pour valider la phylogénèse proposée dans le doc. de référence.
On sait que le partage d’états dérivés permet de retracer l’évolution au sein d’une « collection » d’êtres vivants.
- Les états dérivés « présence de défenses » et « présence de membres postérieurs » sont partagés par les
Eléphants indiens et africains, le Mammouth, le Dinothérium et le Mastodonte mais pas par le Lamantin, ce
qui revient à créer un sous groupe des Proboscidiens au sein du groupe des Téthythériens.
- L’’état dérivé « trou auditif hors de l’os squamosal » caractérisent les éléphants indiens et africains, le
mammouth, et le Mastodonte (mais pas le Dinothérium ni le Lamantin) donc ces 4 espèces possèdent un
AC qu’ils ne partagent pas avec le Dinothérium.
- L’état dérivé « carpes larges et courts » caractérisent les éléphants indiens et africains, le mammouth (mais
pas le Dinothérium ni le Mastodonte ni le Lamantin) donc ces 3 espèces possèdent un AC exclusif
- L’état dérivé « orbites anguleuses » rassemble les éléphants indiens et le mammouth, (mais l’éléphant
africain, le Mastodonte, le Dinothérium ni le Lamantin ne possèdent ce caractère dérivé) donc ces 2
espèces possèdent un AC exclusif
- L’état dérivé « Défenses spiralées » isole le Mammouth des autres espèces.
On peut donc
confirmer l’arbre
phylogénétique
proposé :
1
/
3
100%