Thème : Génétique et évolution

1
Thème : Génétique et évolution
Chapitre 1 : Brassage génétique, innovation génétique et
évolution des génomes
Introduction
Les espèces qui utilisent la reproduction sexuée conservent leurs plan d’organisation au cour des
générations mais se caractérisent aussi par leur très grandes diversité génétique et phénotypique
Les deux mécanismes fondamentaux de la reproduction sexuée sont à l’origine de cette diversité :
La méiose permet la formation des gamètes
La fécondation permet la rencontre des gamètes
Comment ces mécanismes contribuent-il à la diversité génétique des individus ?
I. Méiose et production de gamètes haploïdes variés.
1) La méiose dans le cycle de développement d’un organisme diploïde.
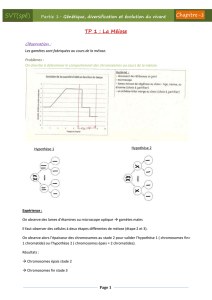
Activité 1 : Découvrir le rôle de la méiose dans le cycle de développement
d’un organisme diploïde.
Bilan cours
a) Place de la méiose dans le cycle de développement des diploïdes.

2
1 : cellule a 2N chromosome monochromatidiens
2 : synthèse d’ADN = duplication (phase S)
3 : cellule a 2N chromosome bichromatidiens (cellule diploïde)
4 : première division de méiose, passage a 2 cellules à N chromosomes bichromatidiens
(cellules haploïde)
5 : seconde division de méiose, passage à 4 cellules a N chromosomes monochromatidiens
(cellules haploïde)
Un organisme diploïde possède des paires de chromosomes, ces cellules sont dites à 2N
chromosome n étant le nombre de paire de chromosome. Dans le cycle de développement dans
organisme diploïde, la méiose est une division qui permet la formation des cellules sexuelles ou
gamètes males et femelles, qui sont haploïdes. La méiose permet de diviser par deux la quantité
Schéma : Le cycle de développement d’un organisme diploïde
Schéma : Quantité d’ADN dans une lignée cellulaire pendant la
méiose

3
d’ADN présente dans les gamètes (voir graphique ci-dessus.) ces gamètes ne possèdent plus de paire
de chromosomes et sont donc des cellules à N chromosomes.
La méiose permet donc d’obtenir l’haploïdie, c'est-à-dire permet le passage d’un état diploïde a un
état haploïde. Chez les diploïdes la méiose précède obligatoirement la fécondation qui reforme de
nouveaux organismes diploïdes. Ceci est indispensable pour le caryotype d’une espèce se maintienne
au cours des générations.
La méiose est obligatoirement précédée par une réplication semi-conservative de l’ADN, qui permet
de doubler la quantité d’ADN et de passer de chromosomes à une chromatide à des chromosomes à
deux chromatides.
b) Les étapes de la méiose
La méiose est composée de successions de deux divisions.
La première division est une division réductionnelle car elle sépare toutes les paires de
chromosomes, c’est donc elle qui permet de passer d’une cellule 2N à une cellule N
Prophase : les chromosomes sont visibles et possèdent deux chromatides chacun. Les
homologues (les chromosomes d’une même paire) s’apparient, ils forment des bivalents ou
tétrades. Au cours de cette étape des échanges de portion de chromatides peuvent se
produire entre deux homologues d’une même paire : se sont les « crossing-over ».
Métaphase : les chromosomes homologues se placent de part et d’autre de la plaque
équatoriale. Contrairement à la mitose ils ne se divisent pas au niveau du centromère.
Anaphase : les chromosomes homologues se séparent et migrent aux pôles de la cellule.
Télophase : deux cellules haploïdes à N chromosome se forment
La seconde division est une division équationnelle qui équivaut à une mitose concernant les deux
cellules à N chromosomes issues de la première division de méiose.
Prophase : les chromosomes à deux chromatides sont bien individualisés
Métaphase : tous les chromosomes se rangent à l’équateur de la cellule et ils se coupent au
niveau du centromère.
Anaphase : deux lots identiques de N chromosomes à une chromatide migrent aux pôles de
la cellule.
Télophase : Quatre cellules à N chromosomes s’individualisent.
Le mécanisme du crossing-over entre deux chromosomes homologues en prophase I de méiose
Début de Prophase I de méiose
(Appariement chromosomes homologues)
Fin de prophase I de méiose
(Formation de chromosomes recombinés)
Crossing-over

4
Conclusion : bilan de la méiose
Exercice d’application page 30 ex 7
1) 4 : prophase I 2Q 2N
3 : métaphase I 2Q 2N
8 : anaphase I2Q 2N
5 : télophase I Q N
2 : Prophase II Q N
6 : Métaphase II Q N
7 : Anaphase II Q N
1 : Télophase II Q/2 N
2) Les brassages génétiques de la méiose et la production de gamètes
haploïdes variés
a) Le brassage intra-chromosomiques

5
P
Génotype → e
Phénotype →
_ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _
Gamètes ↓
↘ Gènes eb+ ; vg+ eb ; vg
F1
Génotype→ eb+//eb ; vg+//vg
Phénotypes de tous les F1 →
2ème croisement
Parent F1 femelle x double récessif mal
Génotype eb+//eb ; vg+// vg eb//eb ; vg//vg
Phénotype
Descendants : à compter !
Gènes : couleurs du corps (2 allèles)
Clair
eb+
Ebène
eb
Gènes : longueur des ailles (2 allèles)
Longues
vg+
Vestigiales
vg
1
2
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
1
/
29
100%