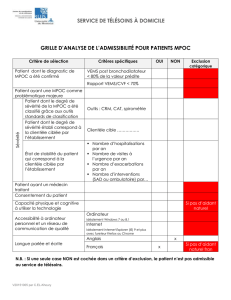
caractérisation des microarn dans les muscles locomoteurs des

CARACTÉRISATION DES MICROARN DANS
LES MUSCLES LOCOMOTEURS DES
PATIENTS ATTEINTS D’UNE MALADIE
PULMONAIRE OBSTRUCTIVE CHRONIQUE
Mémoire
Alexandra Porlier
Maîtrise en médecine expérimentale
Maître ès sciences (M.Sc.)
Québec, Canada
© Alexandra Porlier, 2015


iii
Résumé
L’atteinte des muscles locomoteurs est une manifestation systémique de la
maladie pulmonaire obstructive chronique (MPOC). Ce mémoire a pour objectif
général l’étude des microARN (miR) impliqués 1) dans la myogenèse et 2) dans
l’angiogenèse musculaire squelettique. Tout d’abord, l’expression de miR-1 influence
la myogenèse dans les muscles locomoteurs chez des patients atteints d’une MPOC,
un effet qui ne serait pas médié par IGF-1 ni par HDAC4. Ensuite, nous avons montré
que l’expression de miR-1 semble négativement reliée à l’expression de VEGF dans le
vastus lateralis chez les patients atteints d’une MPOC. L’ensemble de ces résultats
suggère que miR-1 joueraient un rôle important dans les voies de signalisation
impliquée dans la régulation de la myogenèse et de l’angiogenèse des muscles
locomoteurs dans la MPOC.


v
Abstract
Limb muscle dysfunction is a systemic manifestation of COPD. This master’s
thesis has for objective the investigation of microRNAs (miRs) involved 1) in
myogenesis and 2) in skeletal muscle angiogenesis of patients with COPD. First, we
showed that miR-1 appears to influence myogenesis in COPD, but not through IGF-1
or by HDAC4. We also showed that miR-1 negatively influences protein expression of
VEGF in skeletal muscle angiogenesis of patients with COPD. Overall, these results
suggest that miR-1 may play an important role in the regulation of limb muscle
myogenesis and angiogenesis in COPD.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
1
/
133
100%