Le mystère de l`homogénéité génétique de l`humanité - iPubli

NOUVELLES
1995 ; 11 : 771
Le mystère de l'homogénéité génétique
de l'humanité
*
770

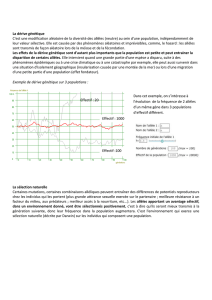
1
S ; :
N,
S ;
771
1
/
2
100%