Correction du DS 8 Thème 1

Correction du DS 8
Thème 1
Document 1.1
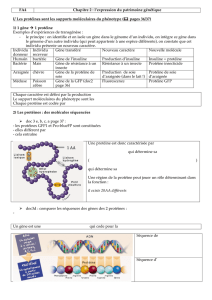
L’immunolocalisation permet, grâce à la spécificité des anticorps, de révéler avec précision la
présence d’une protéine. Ici, les grains d’or sont visibles sur les clichés de microscopie électronique
et sont localisés à l’emplacement e la PEPc. La PEPc se situe dans le cytosol majoritairement!: on
observe néanmoins quelques enzymes dans le stroma du chloroplaste du haut.
Document 1.2
1.2.A - Plante CAM!: le taux d’absorption de CO2 est maximal durant la nuit, avec un taux de 5
µmol.m-2.s-1. Lors de la période lumineuse, le taux d’absorption est nulle, voire même négatif, ce qui
signifie que la plante peut rejeter du CO2 (phénomène de respiration).
Plante en C3!: la fixation n’est réalisée que le jour, avec un taux compris entre 0,2 et 1,1
µmol.m-2.s-1. La nuit, la plante libère du CO2, produit de la respiration cellulaire.
Bilan!: les plantes CAM fixent le CO2 la nuit, avec une efficacité 5 fois supérieure à celle des plantes
en C3.
1.2.B – Le graphique 1 montre que la PEPc a une forte activité (proche de 100%) la nuit (après une
délai d’environ 2h) mais seulement 60% le jour.
Le graphique 2 montre à l’inverse que l’activité enzymatique de la RubisCO est maximale le jour
(au-dessus de 15 µmol CO2 .m-2.s-1 de 9h30 à 18h) mais reste inférieure à 15 µmol.m-2.s-1 la nuit.
Les deux enzymes semblent se relayer dans le temps et donnent l’idée d’une fixation en 2 temps du
CO2 nécessaire à l’autotrophie.
Document 1.3
Le graphique montre que le taux de malate augmente durant la nuit, de 5 à 55 µmol/g de matière
fraîche.
Le malate serait donc le produit de la réaction catalysée par la PEPc!: celle-ci est en effet une PEP
carboxylase, ce qui signifie qu’elle fixe un CO2 sur le PEP.
PEP + CO2 --> malate
Le jour, le taux de malate décroît en quelques heures et devient quasiment nul vers 16h. Le malate
est donc consommé le jour. On peut imaginer que ce malate libère le CO2 fixé la nuit et permet le
fonctionnement de la RubisCO montré sur le document précédent puisque le substrat de la
RubisCO est une molécule de CO2 (et non directement le malate) qui est liée à un ribulose 1-5 diP.
Malate --> CO2 + pyruvate
Le CO2 ainsi libéré permet de réaliser la photosynthèse même si les stomates sont fermés à cause
de la chaleur de la journée et protège donc la plante CAM de la déshydratation.
Schéma bilan

Thème 2
Document 2A1
L’absence de gène cerberus provoque l’absence d’œil et de structures céphaliques. Ce gène est
donc un gène du développement, impliqué dans l’induction de la tête.
L’injection d’ARNm de cerberus en position ventrale postérieure et antérieure induit l’apparition de
têtes surnuméraires à l’endroit même où l’ARNm a été injecté. L’ARNm a été traduit en une protéine
qui a induit une tête entière. Il s’agit donc bien d’un gène architecte!: sa présence a modifié le
devenir des cellules et a induit une cascade d’activation menant à tous les territoires d’une tête.
Bilan!: cerberus est un gène du développement de type «!gène architecte de la tête!».
Document 2A2
Les blastomères dorsovégétatifs du stade 8 cellules constituent le futur endoderme dorsal.
Là où cerberus sera exprimé, le gène LacZ le sera aussi puisqu’il dépend du même promoteur. La
présence du produit de LacZ, la β-galactosidase, est facilement détectable par une réaction
colorée : les territoires qui apparaissent en bleu sont donc ceux qui ont exprimé LacZ et donc
cerberus.
Au stade de gastrulation, on observe que la région qui exprime cerberus correspond à de
l’endoderme dorsal, situé sous l’ectoderme. La couleur bleue est plus intense et large du coté
antérieur de l’embryon!: cela correspond à la zone de la future tête.
Bilan!: cerberus est un gène du développement qui s’exprime dans l’endoderme dorsal au cours de
la gastrulation et induit la formation d’une tête.
Document 2B
On cherche à connaître l’effet de BMP4 sur la formation de la tête et de préciser son interaction
avec cerberus.
CO2
stomates
ouverts
CO2
malate
PEPc
malate
PEP
NUIT
CO2
malate
malate
pyruvate
JOUR
lumière
NADPH,H+
ATP
SUCRE
Calvin

Le gène de BMP4 est associé au promoteur de cerberus donc là où s’exprime cerberus s’exprime
aussi BMP4, en quantité proportionnelle. Plus on a injecté d’ADN chimère, plus la quantité de
BMP4 produit sera importante car il y aura davantage de copies du gène.
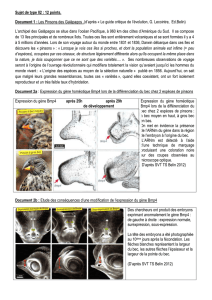
Les clichés D, E et F de bourgeons caudaux montrent que la tête est absente quand cerberus est
en présence d’une grande quantité de BMP4, mais que l’effet de BMP4 s’amenuise quand sa
concentration diminue.
Le témoin présente une tête normale.
BMP4 s’oppose donc à cerberus dans la formation de la tête. Les 2 gènes sont antagonistes.
BMP4 n’agirait pas sur la transcription de cerberus, puisqu’ils sont sous dépendance du même
promoteur, mais pourrait interférer avec l’action de la protéine Cerberus.
Document 2C1
Cas Krox 20!: témoin qui permet de comparer l’intensité des bandes protéiques.
Le profil obtenu est similaire dans les 2 cas, embryon injecté ou non. BMP4 ne semble donc pas
avoir d’influence sur l’expression de Krox20.
Cas de XBF1
Un embryon non injecté (témoin) montre l’apparition de la protéine XBF1 au stade 13, en forte
quantité!: cette présence persiste au moins jusqu’au stade 15. Le stade 11 est celui de la
gastrulation d’après le document 2A2!: le stade 13 pourrait se situer au moment de la neurulation
ou un peu après.
En présence de BMP4, le gène xbf1 n’est pas exprimé!: BMP4 semble réprimer xbf1, responsable
du développement de l’encéphale. Cet effet est cohérent avec l’absence de tête observée en 2B.
Document 2C2
Test des conditions expérimentales
La ligne de myosine montre la présence de la protéine dans les 2 cas!: cela signifie que l’ARNm de
myosine a pu être traduit in vitro!: ce témoin valide l’efficacité du système de traduction.
La traduction de l’ARNm de cerberus est possible en présence de l’ARNm de cerberus et d’un autre
ARN non complémentaire.
Par contre, l’ARN CerMo bloque la traduction!: il se lie par complémentarité à l’ARNm de cerberus,
et bloque, par interférence, la traduction.
L’ARN interférent CerMo est donc efficace. Il peut être utilisé dans la suite comme outil pour
désactiver l’apparition de la protéine Cerberus.
La ligne ODC montre des spots identiques!: les processus de culture, extraction, séparation par
électrophorèse sont donc efficaces. Cette ligne témoin valide l’expérience.
Colonne 1
Le témoin est la colonne St 30/31!: on y observe l’expression de xbf1. Un embryon «!normal!»
possède donc cette protéine au stade 30/31.
Colonne 2
L’ectoderme dorsal seul ne possède pas la protéine XBF1. Le gène n’y est donc pas exprimé.
Colonne 3
L’ectoderme dorsal associé à l’endoderme dorsal présente XBF1!: on peut alors proposer 2
hypothèses!:
-XBF1 est synthétisée et présente dans l’endoderme!;
-XBF1 a été exprimée dans l’ectoderme suite à l’induction par l’endoderme.
La première hypothèse peut être éliminée par la donnée du document 2C!: XBF1 est une protéine
marqueur du télencéphale, de nature ectodermique.

Colonne 4
Si l’endoderme n’a pas de protéine Cerberus (dont la traduction est bloquée par l’interférence de
CerMo), alors l’ectoderme dorsal ne produit pas XBF1.
Cela confirme que l’induction de XBF1 est issu de ADE et précise que cette induction se fait par la
protéine Cerberus.
Bilan
L’endoderme antérodorsal exprime cerberus dès le stade de gastrulation (11). Son expression fait
apparaître la protéine Cerberus dont un effet est l’activation de l’expression du gène xbf1 de
l’ectoderme dorsal, à partir du satde 13 (neurulation!?) conduisant à la mise en place du
télencéphale.
La protéine BMP4 bloque l’effet activateur de xbf1 par Cerberus.
Cerberus a peut-être d’autres effets inducteurs de tête.
cerberus# # # Cerberus# # # xbf1# # # télencéphale
# expression dans# # # # ectoderme
# l’endoderme
# # # # # # BMP4
########### TETE
1
/
4
100%