Document

REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTERE DE L’ENSEIGNEMENT SUPERIEUR ET DE LA
RECHERCHE SCIENTIFIQUE
UNIVERSITE EL-HADJ LAKHDAR BATNA
FACULTE DES SCIENCES
DEPARTEMENT DES SCIENCES DE LA NATURE ET DE LA VIE
MÉMOIRE
Présenté
pour l'obtention du diplôme de
MAGISTER
Filière
Biologie
Option
Biologie animale
Présenté par :
FRIHA SAMIRA
Thème
Étude des variations cycliques de l’activité enzymatique et
électrolytique dans les sécrétions intra utérines et oviductaires au
cours d’un cycle œstral de la vache
Devant le jury :
Présidente : Mme Hambaba L. Professeur. Université El-Hadj Lakhdar Batna
Rapporteur : Mr Yahia M. Professeur. Université El-Hadj Lakhdar Batna
Examinateur : Mr Kaidi R. Professeur. Université Saâd D’Ahlab Blida
Examinateur : Mr Laroui S. Professeur. Université El-Hadj Lakhdar Batna
Année universitaire : 2011-2012

Remerciements
Au nom de DIEU, Omnipotent, Omniscient, qui m’a guidé pour réaliser
ce travail.
Je tiens à remercier vivement le docteur : Yahia M; Professeur
à L’Université de Batna et lui témoigne ma profonde
reconnaissance, pour la proposition du thème de mon mémoire et pour la
direction parcimonieuse de mes travaux en dépit des multiples occupations
que lui imposent l’enseignement et la recherche tout particulièrement.
Puis, j’exprime ma profonde gratitude au Docteur Yahia Atia; pour
l’aide précieuse qu’il m’a apporté.
A Mme Hambaba L. Professeur à l’université El-Hadj Lakhdar Batna
Qui nous a fait l'honneur d'accepter la présidence du jury de notre mémoire.
Hommages respectueux.
A Monsieur le Professeur Laroui S Professeur à la faculté de médecine de Batna
Qui nous a fait l'honneur de participer à notre jury du mémoire,
Sincères remerciements et profonde reconnaissance.
A Monsieur Kaidi R, Professeur à l’université de Blida,
Malgré vos multiples occupations, vous avez accepté de siéger dans notre jury,
Recevez notre respectueuse considération.
Par ailleurs, je suis redevable à tous qui m’ont aidé.

Dédicace
A mes parents
A mes sœurs
A mon frère
A Ramdhane, une tendre pensée
A toute ma famille.
A mes amis,
A tous ceux qui, de près ou de loin, ont participé à la réalisation de ce travail.

Table des matières
Liste des abréviations
Liste des figures
Liste des tableaux
Introduction
Partie théorique
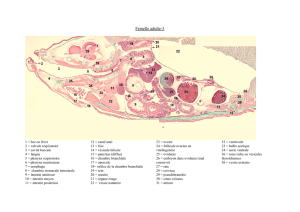
Chapitre I. Rappels anatomo-histologiques du tractus génital de la vache
I.1 Anatomie du système reproducteur de la vache...........................................................1
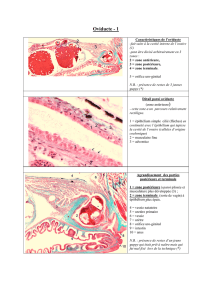
I.2 Histologie du système reproducteur de la vache ..........................................................5
Chapitre II. Rappels physiologiques du tractus génital de la vache
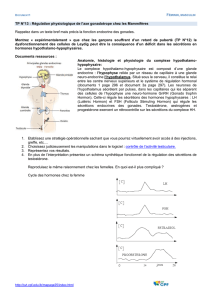
II.1 Cycle œstral bovin ....................................................................................................9
II.1.1 Notions de vague folliculaire, recrutement, sélection et dominance ...............11
II.1.2 Régulation hormonale du cycle sexuel...........................................................13
II.2 Caractéristiques physiologiques du système reproducteur de la vache .......................15
II.2.1 Caractéristiques physiologiques de l’oviducte ...............................................15
II.2.2 Caractéristiques physiologiques de l’utérus ...................................................18
II.3 Rôles physiologiques du système reproducteur de la vache ......................................19
II.3.1 Rôles physiologiques de l’oviducte ..............................................................19
II.3.2 Rôles physiologiques de l’utérus ...................................................................22
Chapitre III. Propriétés biochimiques des sécrétions génitales de la vache
III.I Sécrétions tubaires ...................................................................................................25
III.1.1 Origine du fluide de l’oviducte.....................................................................25
III.1.2 Variations au cours du cycle ........................................................................25
III.1.3 Caractéristiques biochimiques des sécrétions oviductaires ........................... 25

III.2 Sécrétions utérines ...................................................................................................28
III.2.1 Origine du fluide utérin ................................................................................28
III.2.2 Variations au cours du cycle ........................................................................28
III.2.3 Caractéristiques biochimiques des sécrétions utérines .................................28
Partie expérimentale
Chapitre I. Matériel et méthodes
I. 1 Présentation de l’exploitation ....................................................................................35
I.2 Protocole expérimental ...............................................................................................35
I. 3 Analyses statistiques ..................................................................................................41
Chapitre II. Résultats et discussion
II.1 Electrolytes dans le sérum et les sécrétions génitales de la vache ...............................42
II.2 Propriétés enzymatiques des sécrétions endométriales ...............................................55
II.3 Protéines totales dans le sérum et dans les sécrétions génitales ..................................59
Conclusion
Bibliographie
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
1
/
100
100%