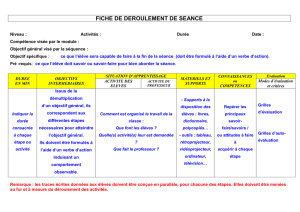
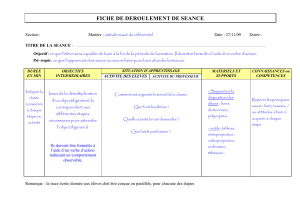
La regression du mesonephros chez Fembryon de

J. Embryol. exp. Morph., Vol. 15, 3, pp. 397-419, June 1966 397
With
11
plates
Printed in
Great
Britain
La regression du mesonephros chez
Fembryon de poulet
Etude des activites de la phosphatase acide et des cathepsines.
Analyse biochimique, histochimique et observations
au microscope electronique
Par BERTHE SALZGEBER1
Institut d'Embryologie et de Teratologie Experimentales du CNRS,
Nogent-sur-Marne {Directeur: Professeur Et. Wolff)
& RUDOLF WEBER2
Institut de Zoologie, Division de Biologie Cellulaire
et d'Embryologie chimique, Berne
INTRODUCTION
Au cours du developpement des vertebres, on observe de nombreux exemples
de regression spontanee de tissus ou meme d'organes (Gliicksmann, 1951).
Ainsi le canal de Miiller de l'embryon $ de poulet represente un exemple
typique d'atrophie d'un organe au cours de la vie embryonnaire
(Wolff,
1953)
et le determinisme de cette regression a fait l'objet de nombreux travaux
(Wolff,
1936;
Wolff,
Lutz-Ostertag & Haffen, 1952; Lutz-Ostertag, 1954). Un
autre exemple est celui de la resorption de la queue des tetards d'amphibiens
anoures au cours de la metamorphose (Kendall, 1919; Vulpian, 1859; Weber,
1962,
1963a).
Des etudes biochimiques ont permis de completer les donnees morphologiques
et ont contribue a eclaircir le mecanisme de la regression de ces organes. Citons
les travaux de Scheib-Pfleger (1955), Brachet, Decroly-Briers & Hoyez (1958) et
Scheib-Pfleger & Wattiaux (1962) sur la regression des canaux de Miiller de
l'embryon $ de poulet et les travaux de Weber (1957a, b), Weber & Niehus
(1961),
Weber (19636) et Eeckhout (1964) sur l'atrophie de la queue de tetards
d'amphibiens anoures. L'ensemble de ces recherches demontre que l'involution
des tissus coincide avec une augmentation significative de certaines hydrolases
1
Adresse
de
Vauteur:
Institut d'Embryologie et de Teratologie Experimentales du Centre
National de la Recherche Scientifique, Nogent-sur-Marne, France.
2
Adresse de
Vauteur:
Institut de Zoologie, Division
de
Biologie Cellulaire et d'Embryologie
chimique, Berne, Suisse.

398 B. SALZGEBER & R. WEBER
acides. II convient de noter que ces travaux ont ete inspires en grande partie
par les recherches biochimiques de de Duve et collaborateurs (de Duve et al.
1955) sur la distribution intracellulaire d'enzymes dans le foie de rat. Us ont
abouti a la decouverte d'une nouvelle categorie de granules cytoplasmiques
appeles 'lysosomes'. Les auteurs ont demontre que ces organites contiennent
un assortiment specifique d'hydrolases acides. Liberees des granules, ces en-
zymes sont capables d'agir sur les principaux constituants cellulaires. Pour
de Duve (1958), les lysosomes sont les instruments chimiques de l'autophagie
cellulaire. Celle-ci serait declenchee par la liberation des hydrolases acides des
lysosomes, ce qui devrait expliquer l'augmentation de l'activite de ces enzymes
dans les tissus en voie de regression.
Les techniques biochimiques ayant permis de mettre en evidence des activites
enzymatiques, mais non leur localisation dans le tissu in situ, des chercheurs se
sont adresses recemment aux techniques d'histochimie et de microscopie electro-
nique. Dans les queues de Xenopus en regression, Salzmann & Weber (1963) et
Weber (1964) observent de nombreux macrophages riches en phosphatase
acide et en esterases de type cathepsine, alors que ces elements sont peu nom-
breux et peu actifs au stade precedant la metamorphose.
Ces auteurs pensent a une activation locale des macrophages dans le tissu
en regression et attribuent l'augmentation de l'activite de ces enzymes a la
neosynthese d'enzymes par les cellules phagocytaires. Dans le canal de Muller de
l'embryon de poulet S, Scheib (1964) signale egalement unenette augmentation
de l'intensite de la reaction de la phosphatase acide au cours de la regression
de l'organe. De nombreuses granulations sont visibles du cote apical des cellules
epitheliales et dans le tissu
conjonctif,
en particulier dans les cellules de type
macrophage.
Le mesonephros de l'embryon de poulet represente un autre cas d'atrophie
spontanee d'un organe
(Wolff,
1953). Au cours de son developpement embryon-
naire, le mesonephros croit, augmente de poids pour atteindre un developpe-
ment maximum au stade de 14 jours, puis il entre dans une phase de regression
caracterisee par la diminution de la taille et la perte de poids de l'organe
(Firket, 1920; Atwell & Hanan, 1926; Schmalhausen, 1927; Stampfli, 1950;
Haffen, 1951a;
Romanoff,
1960; Voile & Beaumont, 1964). Les etudes histolo-
giques montrent differents aspects de la degenerescence: alteration des glomerules
de Malpighi, involution des tubes mesonephretiques qui se retrecissent et se
transforment en cordons pleins, destruction de cellules qui se detachent et
tombent dans la lumiere des tubes.
Des dosages d'azote total effectues par Haffen (1951Z?) ont montre que la
diminution du poids de l'organe entre le 15e et le 21e jour de l'incubation
s'accompagne d'une perte en matiere azotee. Une 2e etape dans l'etude de la
regression du mesonephros consiste dans l'analyse biochimique de l'activite de
certaines enzymes dans l'organe en croissance et en regression. Ainsi l'activite
de l'hypoxanthine deshydrogenase, enzyme jouant un role important dans la

Regression du mesonephros 399
synthese de l'acide urique, augmente pendant le stade de croissance de l'organe
puis diminue au cours des derniers jours de l'incubation (Chaube, 1962;
Croisille, 1962
a).
Effectuant Panalyse immunochimique du rein, Croisille
(1962&) a d'autre part montre la presence d'importantes activites esterasiques
dans les extraits de mesonephros et observe l'apparition entre le 10° et le
12e
jour de l'incubation d'une esterase, specifiquement liee au rein, qui disparait
au moment ou le mesonephros entre dans sa phase de regression. Les enzymes
sont alors presentes dans le metanephros.
Junqueira (1952) et Russo-Caia (1962) ont etudie le comportement des
phosphomonoesterases acides et alcalines au cours du developpement du
systeme renal de l'embryon de poulet, au moyen de methodes quantitatives et
histochimiques. Pour la phosphatase acide, les resultats de ces auteurs different
quant a la duree de l'augmentation de l'activite enzymatique. Celle-ci, selon
Russo-Caia (1962) persiste encore pendant la regression du mesonephros, tandis
que Junqueira (1952) la constate seulement durant la croissance et signale une
diminution pendant Finvolution de cet organe. Ces resultats ne permettent pas
de tirer des conclusions precises sur le role de la phosphatase acide dans la
regression du mesonephros, d'autant plus que ces auteurs n'ont pas aborde le
probleme de la solubilisation de cette enzyme. Tout recemment, Tilney (1964)
a mis en evidence a l'aide du microscope electronique la localisation de la
phosphatase acide dans des elements cytoplasmiques du type 'corps denses',
qui apparaissent nombreux au stade de l'eclosion.
Notre travail a pour but de preciser le comportement des hydrolases acides
dans un cas modele d'autolyse cellulaire non liee a Faction des hormones. Nous
nous sommes contentes d'etudier deux enzymes typiques des lysosomes: les
cathepsines et la phosphatase acide, en comparant les activites et l'etat de ces
enzymes dans le mesonephros du
14
e
jour (croissance) et du
21e
jour (regression).
Les etudes biochimiques ont ete completees par des recherches histochimiques
et par des observations faites au microscope electronique.
MATERIEL ET METHODES
(1) Techniques biochimiques
Les embryons de poulet ages de 14 jours (stade de croissance) et 21 jours
(stade de regression) sont disseques dans du liquide de Ringer froid. Les
mesonephros sont preleves et immerges dans du saccharose
0,25
M
glace.
Pour chaque experience qui comprend une determination de l'activite des
cathepsines, de la phosphatase acide et de l'azote total, on utilise 2 ou 3 em-
bryons, soit 4 ou 6 mesonephros selon le stade de prelevement de ces organes.
Apres prelevement, on homogeneise les organes avec une solution glacee de
saccharose 0,25
M
+ versene 10~
3
M, de facon a obtenir un homogenat de con-
centration 10%. On determine ensuite par pesee le volume de Phomogenat
ainsi obtenu. Les activites enzymatiques sont rapportees: soit a l'unite d'azote

400 B. SALZGEBER & R. WEBER
total utilisee pour le test (activite specifique) soit a la quantite d'azote total de
l'organe (activite totale).
L'activite des cathepsines est determinee selon la methode colorimetrique
de Duspiva (1939) modifiee par Von Hahn & Herrmann (1962). A 50/^1 de
substrat, contenant 1% de caseine et 36% d'uree, ajuste a pH 5,0 par le
tampon Mcllvaine, on ajoute
10
y\ d'homogenat a 1,25 ou 2,5 %. Apres 2 heures
d'incubation a 37 °C, le melange est precipite par 60
jul
d'acide trichloracetique
10%
et conserve pendant 12 heures a basse temperature (0-4 °C). Apres centri-
fugation,
100 {A
de surnageant sont recueillis et traites par le reactif de Folin-
Ciocalteu; l'intensite de la couleur est mesuree au spectrophotometre Zeiss
PMQ II a 780 m/*. La concentration de substances 'Folin' positives a ete
determinee parallelement sur des echantillons temoins non incubes. En retran-
chant ces valeurs temoins des mesures d'extinction d'echantillons incubes, on
obtient la valeur d'extinction qui correspond reellement a l'activite enzymatique
de l'homogenat.
Pour le dosage de la phosphatase acide, nous avons suivi la methode decrite
par Weber & Niehus (1961).
30
fi\ de substrat, contenant 0,028 M de /?-glycero-
phosphate de sodium dans un tampon acetate a pH 5,0, sont incubes pendant
30min. a 37° avec 30
[A
d'homogenat a 0,5 et 1,0% en milieu hypotonique.
Le melange est ensuite precipite avec 60/*1 d'acide trichloracetique a 5%.
Apres centrifugation, on recueille
100
/d du surnageant. Les phosphates liberes
dans le surnageant sont doses d'apres la methode colorimetrique de Kuttner &
Cohen (1927). La mesure des valeurs d'extinction est effectuee a 710 m/i. Des
temoins non incubes ont servi a la determination de la teneur en phosphate libre
dans l'homogenat. Celle-ci sert de correction pour l'obtention des valeurs
d'extinction des echantillons incubes.
Les determinations d'enzymes ont ete realisees dans des conditions telles
que le phenomene de latence soit exclu, soit par l'utilisation d'un milieu hypo-
tonique (phosphatase acide) soit par l'emploi de forte concentration d'uree
provoquant une solubilisation des proteines (cathepsines).
Afin de comparer le degre de solubilisation des enzymes dans les tissus en
croissance et en regression (cf. p. 406) des fractionnements ont ete effectues
dans des conditions iso- et /*y/?otoniques.
L'azote total qui permet d'evaluer la teneur en tissus des homogenats est
dose selon une ultramicromethode de type Kjeldahl mise au point par Boell &
Shen (1954). Quelques dosages d'azote proteinique ont ete effectues selon la
methode de Von Hahn & Herrmann (1962).
(2) Techniques histochimiques
Les organes sont fixes pendant 16 heures a 0 °C dans une solution de formol
a 4% neutralisee avec CaCO3 en presence de saccharose a 10%. Apres lavage
dans l'eau courante, les pieces sont incluses dans du muscle de Rat et immediate-
ment congelees a l'azote liquide (15 a 20 secondes). Les sections effectuees au
cryostat (-10 °C) sont recueillies sur lamelles.

Regression du mesonephros 401
Dans le cas de la phosphatase acide, la localisation de l'activite enzymatique
a ete demontree selon les methodes de Burstone (1958) et de Barka &
Anderson (1962).
Le substrat utilise est le naphtol AS-TR-Phosphate. Le naphtol libere par
l'hydrolyse enzymatique est visualise par couplage avec un sel de diazonium
(FastDark Blue RapH 5,2 selon Burstone (1958) ou rhexazonium pararosaniline
a pH 5,0 (Barka & Anderson, 1962).
Les esterases non specifiques sont mises en evidence selon la methode de
Holt (1958) avec l'o-acetyl-5-bromoindoxyle a pH 7,0 comme substrat.
(3) Etudes au microscope electronique
Des fragments de mesonephros, stades de croissance (12 jours) et stades de
regression (21 jours, 2e et 7e jour apres l'eclosion) sont fixes a l'acide osmique
1 % dans le tampon de Michaelis a pH 7,2 et en presence de
10
% de saccharose.
Apres deshydratation par l'acetone, les tissus sont inclus soit dans PAraldite, le
Vestopal W ou le Durcupan AMC. Les pieces sont coupees a l'aide d'un
ultramicrotome Porter-Blum. Les coupes sont colorees par une solution de
citrate de Pb (Reynolds, 1963) et observees au microscope electronique Siemens
Elmiskop I.
Les lieux d'activite de la phosphatase acide sont mis en evidence a l'aide de la
methode de Miller (1962) utilisant la technique de Gomori.
RESULTATS
(A) Evolution de Vazote total
Les recherches confirment les travaux de Haffen (19516) et montrent que
l'involution du tissu se manifeste par une perte considerable de la substance
azotee (Tableau 1). Les pourcentages de perte de poids frais et d'azote total entre
le 14e et le
21e
jour de l'incubation sont semblables et atteignent respectivement
58,6%
et 62,2%.\
Tableau 1. Perte en azote total et en poids fra\s dans le mesonephros
*
Poids frais par Azote total par
mesonephros mesonephros
(mg)
Stades (x±sz) N (x±sz) N
14
j.
12,7910,65! 10 228,2± 10,13 8
21
j.
5,310,36! 10
86,13
±7,90 8
Perte en % du 14e 58,6% — 62,2% —
au 21ejour
1 Valeurs approximatives (liquide adherent).
x = moyenne, s^ = erreur standard de la moyenne, N = nombre de determinations.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
1
/
35
100%