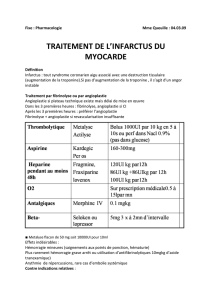
Physiologie de la fibrinolyse

Référence document : Phy3
Rédigé par : Professeur Jean-François
ABGRALL
Indice
0 Page 1 de 1
Titre du document : Physiologie de la fibrinolyse
Physiologie de la fibrinolyse
Professeur Jean-François ABGRALL


Référence document : Phy3
Rédigé par : Professeur Jean-François
ABGRALL
Indice
0 Page 3 de 3
Titre du document : Physiologie de la fibrinolyse
1. Introduction
La fibrinolyse est la dégradation de la fibrine
Deux sites de fibrinolyse
* Fibrinolyse sanguine :
o 1 cellulaire : 80-90% de l'activité
o 2 plasmatique : 20-30% de l'activité
* Fibrinolyse tissulaire :
o réparation tissulaire, lactation
o spermatogénèse
o implantation trophoblaste
Deux sortes de fibrinolyse
* dépendante de la plasmine
* indépendante de la plasmine
Deux étapes :
* transformation du plasminogène en plasmine
* dégradation des substrats par la plasmine
2. Schéma de la Fibrinolyse
+
tPA
+
uPA + -
-
-
PAI
-
A2antiplasmine
lyse de fibrine
Plasmine
Plasminogène
PK + FXII

Référence document : Phy3
Rédigé par : Professeur Jean-François
ABGRALL
Indice
0 Page 4 de 4
Titre du document : Physiologie de la fibrinolyse
3. La fibrinolyse plasmatique
a lieu à la surface de la fibrine
3.1. Plasminogène et plasmine
3.1.1. Plasminogène
précurseur inactif de la plasmine
GP simple chaîne, PM 92 000, 791 AA, 24 ponts S-S, en N terminal : 5 boucles de 80 AA, les kringles, forte affinité
pour la lysine (LBS) = fixation à fibrine, a2-AP, à HRGP, à thrombospondine, à acide e-aminocaproïque.
plasma : 1,5-2 mM/l, augmente lors de infection, inflammation, grossesse.
Plg natif : acide glutamique en NH2 terminal ="Glu-Plg" converti en "Lys-plg" par protéolyse limitée, Lys, val, ou Méth
en NH2 terminal puis Lys-Plg , et libération de peptides de 8 kDa. Lys-Plg a une affinité plus grande pour la fibrine que
Glu-Plg, et une demi-vie plus courte (0,8 vs 2,5 j).
dans le plasma: hydrolyse directe de Glu-plg en plasmine
sur kringles 1,2 et 5 : LBS se lient à fibrine ou à cellule, l'affinité augmente quand la fibrine est dégradée.
3.1.2. Plasmine
sérine-protéase
Glu-Plg est hydrolysé en plasmine (Glu-plasmine) par clivage Arg561-Val562
2 chaînes réunies par ponts S-S, PM : 85 kDa,
o chaîne lourde A (60 kDa) porte les LBS
o chaîne légère B (26 kDa) porte le site actif His603,Asp646,Ser741
La Lys-plasmine ne semble pas exister in vivo.
La plasmine est liée à la fibrine, ce qui augmente son activité
La plasmine dégrade la fibrine, le fg, les facteurs V, VIII, vWF, XIIIa, éléments du complément, la matrice
extracellulaire.
3.1.3. Récepteurs du Plasminogène
existent sur cellule endothéliale, globules blancs, plaquettes, hépatocytes
deux types de récepteurs :
o a-énolase
o gangliosides

Référence document : Phy3
Rédigé par : Professeur Jean-François
ABGRALL
Indice
0 Page 5 de 5
Titre du document : Physiologie de la fibrinolyse
très forte densité > 107 sites par cellule
liaison inhibée par lysine et analogues de lysine (acide tranéxamique)
o gangliosides : lient directement Plg par LBS
o protéines avec lysine en C-terminale et une lysine en N-terminale ( 17 protéines humaines répondent à ces 2
conditions, dont l' a-énolase, protéine intracellulaire de la glycolyse exprimée à la surface.
3.2. Activateurs du plasminogène
3.2.1. t-PA
principalement synthétisé par la cellule endothéliale, produit aussi par cellule mésothéliale, MK, monocyte
gène : chromosome 8
70 pM/ml, demi-vie = 3-6 secondes, à 80 % lié à PAI-1
sérine-protéase, une chaîne, 527 AA, PM 70 kDa
deux formes également actives : sct-PA et tct-PA (protéolyse en Arg275-Ile276 par la plasmine, le Xa, ou la
kallicréine)/p
4 domaines :
o domaine F : résidus 4 à 50 en NH2-terminal, homologue du finger : domaine de fibronectine--> affinité pour
fibrine et pour cellules
o domaine E : 50 à 87 , homologue EGF --> liaison sur cellules
o domaine K1: 87 à 176 ; K2 : 176 à 262 --> liaison à la fibrine
o domaine sérine-protéase : 276 à 527 , site actif His322,Asp371, Ser478
liaison à PAI-1 : finger, K2, et portion terminale
enzyme peu active en l'absence de fibrine, la liaison à la fibrine augmente l'activation du Plg (2 degrés de magnitude =
100 fois)
Le tPA est éliminé par le foie en quelques minutes (tPA/PAI-1 se lie au récepteur LRP/a2-MR de l'hépatocyte).
La plasmine formée à la surface de la fibrine a ses LBS et son site actif occupés, donc est lentement inactivée par a2-
AP (demi-vie 10 à 100 sec).
La plasmine libre est inactivée en 0,1 sec par a2-AP.
La plasmine digère la fibrine, ce qui augmente le nombre de sites lysine, donc augmente la fixation de Plg et l'action
de la plasmine.
3.2.2. u-PA ou Urokinase
rôle secondaire dans la circulation, mais rôle important dans la matrice extracellulaire.
Scu-PA : PM 54 000, 411 AA, chromosome 10
hydrolyse limitée par plasmine ou kallicréine Lys158-Ile159 = tcu-PA ou urokinase
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%