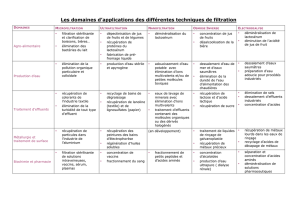
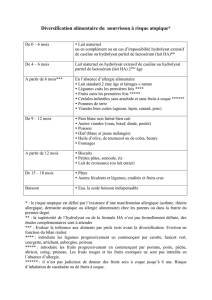
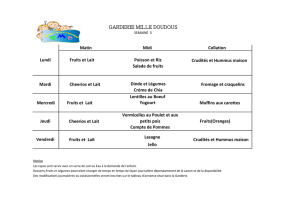
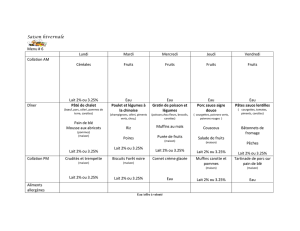
aptitude d`une souche lactique a croitre sur un milieu de culture a

REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE MINISTERE DE
L’ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE SCIENTIFIQUE
Université KASDI MERBAH Ouargla
FACULTE DES SCIENCES DE LA NATURE ET DE LA VIE
DEPARTEMENT DES SCIENCES BIOLOGIQUE
Mémoire
MASTER ACADEMIQUE
Domaine: Sciences de la Nature et de la Vie
Filière: Biologie
Spécialité: Microbiologie Appliquée
Présenté par: Khacef Lynda
Thème
Soutenu publiquement
Le : 14 / 06 /2015
Devant le jury :
Année universitaire 2014-2015
APTITUDE D’UNE SOUCHE LACTIQUE A CROITRE SUR
UN MILIEU DE CULTURE A BASE DE LACTOSERUM
Président SIBOUKEUR OEK Prof. U.K.M.Ouargla
Encadreur Melle SOUID Wafa ( M.A.A) U.K.M.Ouargla
Co-Encadreur Mme BOUDJENAH Saliha (M. C. A) U.K.M.Ouargla
Examinateur Mr BOURICHA ( M.A.A) U.K.M.Ouargla

Remerciements
Avant tout, je remercie Dieu tout puissant de m’avoir accordé la force,
le courage et les moyens afin de pouvoir accomplir ce modeste travail
.
Tous mes vifs remerciements et ma profonde reconnaissance s'adressent
à ma promotrice Mademoiselle SOUID Wafa, MA.A à l’université
KASDI MERBAH Ouargla département des Sciences biologique
et ma co-promotrice Madame
BOUDJENAH-HAROUN Saliha , Maître de conférences A à
l’université K .M . Ouargla
pour leur précieuse aides, leurs conseils, tout au long de l’élaboration de ce
modeste travail.
Je remercie particulièrement
Madame SIBOUKEUR OUMELKHEIR
professeur à l’université KASDI MERBAH Ouargla
pour avoir accepté de présider le jury
Je remercie très
particulièrement
Mr BOURICHA
pour m’avoir fait
l’honneur d’examiner et juger mon travail
Je remercie mes très chers parents pour leur soutien
et leur encouragement
Je remercie particulièrement mon mari Mr HAMMOUM Rachid
pour ses aides, ses
soutiens
, tout au long de l’élaboration de ce
modeste travail.
Mes vifs
remerciements
vont également à Mr EL OUANI Safi le
directeur de la laiterie de Ghardaïa.
Mes
remerciements
vont également à
tous les membres du
laboratoire de microbiologie qui m’ont soutenu.
Enfin je remercie toute personne ayant participé d’une manière ou d’’une
autre à la réalisation de ce modeste travail

Dédicaces
Je dédie ce mémoire à :
Mes très chers parents
Mon adorable mari HAMOUM Rachid qui
n’arrête pas de me soutenir
Mes très chers enfants MOHAMED Arezki et Ritadje
Mon cher frère Djilali.
Mes très chères sœurs Nadia, Fatiha, Hassina, Razika,
Sabrina, Zhira, Froudja, ainsi leurs maris
La mémoire de mes grands parents, mes grandes mères
Mes voisins, mes amis particulièrement
AMINA et mes collègues de travail.
Enfin à tous mes amis qui me sont chers.
Sans oublier la promotion microbiologie 2015
Lynda

Résumé
Dans ce travail, nous nous sommes intéressés à la valorisation du lactosérum. Ce sous
produit des industries laitières, constitue, d’une part un facteur de pollution redoutable et
d’autre part, il renferme des constituants (lactose, éléments minéraux et protéines) ce qui en
fait un excellent milieu de culture pour les micro-organismes. L’objectif de notre travail
consiste à étudier l’aptitude d’une souche lactique (du genre Lactococcus) à croitre dans des
milieux de culture à base de lactosérums doux (bovin et camelin) en utilisant un milieu de
culture M17 comme témoin. Lors de cette étude nous avons isolé à partir du lait camelin
local une souche lactique S1 Gram + en forme de cocci et catalase - qui a été purifiée par la
méthode de stries sur milieu M17. L’identification phénotypique nous a permet de classer
cette souche dans l'espèce : Lactococcus lactis subsp. cremoris.
Nous avons par la
suite
testé l’aptitude de cette souche à croitre dans un milieu de culture à base de lactosérum
bovin et camelin déprotéiné et non supplémenté. Les résultats obtenus ont révélé que les
lactosérums bovin et camelin constituent un milieu de culture adéquat pour la croissance de
cette souche avec une meilleure croissance dans le lactosérum camelin. En effet après 48h et à
pH initial de 6.8 ; une acidification du lactosérum camelin a été considérable (pH 3.97) par
rapport au lactosérum bovin (pH 4.20). La biomasse produite dans le lactosérum camelin
était de 1.50g/1 alors que celle produite dans le lactosérum bovin était de 1g/l.
Mots clé : Aptitude, Lactosérumcamelin, lactosérum bovin, milieu de culture, valorisation,
bactéries lactiques, industrie laitière.

Summary
In this work, we focused on enhancing the whey. This by-product of the dairy industry is
on one hand a formidable source of pollution and secondly, it contains components (lactose,
minerals and protein elements) which make it an excellent culture medium for
microorganisms. The aim of our work is to investigate the ability of lactic strain (genus
Lactococcus) to grow in culture media containing sweet whey (cattle and camels) using an
M17 culture medium as a control. In this study we isolated from the local camel milk lactic
strain S1 Gram-shaped cocci and catalase - that was purified by the method of streaks on M17
medium. Phenotypic identification has allows us to classify this strain in the species
Lactococcus lactis subsp. cremoris. We subsequently tested the ability of this strain to grow in
a bovine whey-based growth medium and camels and not protein-supplemented. The results
showed that bovine whey and camel are an appropriate culture medium for the growth of this
strain with better growth in the camel whey. Indeed after 48 hours and initial pH of 6.8;
acidification of camel whey was considerable (pH 3.97) compared to bovine whey (pH 4.20).
The biomass produced in the camel whey was 1.50g / 1, while that produced in the bovine
whey was 1 g / l.
Keywords: Fitness, Whey camel, bovine whey, culture medium, recovery, lactic acid
bacteria, dairy industry.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
1
/
76
100%