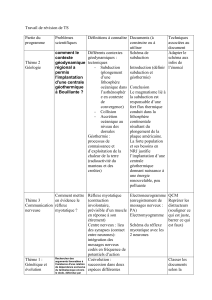
Correction des sujets de bac proposés dans le manuel - Hachette

© Hatier, 2003 1
Correction des sujets de bac proposés dans le manuel
SUJET 1
PARTIE 1 : Les anticorps anti-VIH
Introduction
Une personne est dite séropositive au VIH lorsqu’on a détecté dans son plasma, l’existence
d’anticorps anti-VIH.
La détection de la séropositivité s’effectue grâce à un test Elisa et au Western blot qui
reposent tous les deux sur la formation de complexes antigène – anticorps visualisés par une
réaction colorée. Le Western blot, plus élaboré que le test Elisa, révèle la présence de
plusieurs sortes d’anticorps, chacun réagissant avec une protéine virale définie. Cela traduit la
spécificité de chaque anticorps. Nous allons envisager les caractéristiques de la structure des
anticorps rendant compte de leur capacité à former des complexes antigène – anticorps et de
la spécificité de cette reconnaissance. Nous étudierons enfin, le rôle que jouent ces anticorps
vis à vis de l’infection par le VIH.
I. La structure des anticorps anti-VIH
• Les anticorps anti-VIH n’ont rien de particulier : ils ont la même structure globale que
celle de tout anticorps vis-à-vis d’un antigène . Une molécule d’anticorps est constitué de
quatre chaînes polypeptidiques identiques deux à deux (figure 1 document 2 page 139 du
manuel Tle S) : deux chaînes dites lourdes (H) et deux chaînes dites légères (L).
• La comparaison des séquences d’acides aminés des chaînes lourdes des divers anticorps
anti-VIH révèlent qu’elles possèdent une région constante et une région variable (figure
1). L’expression région constante signifie que la séquence d’acides aminés de cette région
est la même pour toutes les chaînes lourdes de tous les anticorps. En revanche, il existe
des différences dans celle des régions variables de ces chaînes lourdes des divers
anticorps.
De même, la comparaison des chaînes légères des divers anticorps révèle une région
constante et une région variable. Au sein de la région variable, aussi bien des chaînes

© Hatier, 2003 2
lourdes que des chaînes légères, on reconnaît des régions hypervariables où la séquence
d’acides aminés est toujours différente d’un anticorps à l’autre.
• La capacité des anticorps à former des complexes avec des antigènes spécifiques est liée
aux régions variables des chaînes polypeptidiques de la molécule. Plus précisément, les
régions hypervariables d’une chaîne lourde et d’une chaîne variable forment dans
l’espace, un site de reconnaissance d’un antigène précis. Les acides aminés de ce site se
lient spécifiquement à une région de l’antigène (déterminant antigénique). Les sites de
reconnaissance d’un anticorps anti-gp 120 reconnaissent la protéine d’enveloppe gp 120
du VIH, ceux d’un anticorps anti-p 24 reconnaissent la protéine p24 du VIH, etc…
• Chaque molécule d’anticorps possède donc deux sites de reconnaissance de l’antigène.
Elle peut se lier à un molécule d’antigène par un site et à une autre molécule d’antigène
(identique) par le second site : il en résulte qu’un ensemble de molécules d’anticorps et
d’antigènes peuvent former des complexes.
II. Rôle protecteur des anticorps anti-VIH
• En se liant aux protéines de l’enveloppe du virus, les anticorps empêchent la fixation de
celui-ci aux molécules CD4 membranaires des lymphocytes T4. En neutralisant le virus,
les anticorps limitent l’infection des lymphocytes T4.
• La région constante des chaînes lourdes d’anticorps a des propriétés effectrices. Elle peut,
notamment se lier à des récepteurs membranaires des cellules phagocytaires. Les
complexes anticorps anti-VIH – virus peuvent ainsi être phagocytés par ces cellules ce qui
contribue à la réduction du nombre de particules virales (charge virale) dans le milieu
extracellulaire (plasma, lymphe).
Conclusion
Les anticorps anti-VIH sont des molécules effectrices résultant des réactions immunitaires
entraînées par l’infection par le VIH.
La spécificité des complexes qu’ils forment avec les protéines des VIH est liée à l’existence
de deux sites de reconnaissance spécifiques d’un antigène par chaque molécule d’anticorps.
Le rôle protecteur des anticorps anti-VIH vis-à-vis de l’infection est modeste. Comme tous les
anticorps, ils n’agissent que dans le milieu extracellulaire et sont donc sans action sur la
production de nouveaux virions par les lymphocytes T4 infectés.

© Hatier, 2003 3
PARTIE 2 : L’age de deux granites (question 2A)
Il existe plusieurs méthodes de datation absolue des roches basées sur la désintégration
d’éléments radioactifs. Les légendes des axes des abscisses et des ordonnées du graphe
proposé indiquent que ces granites ont été datés par la méthode rubidium – strontium bien
adaptée aux roches magmatiques dont la formation remonte à des centaines de millions
d’années.
Pour chaque minéral du granite, le rapport 87Sr / 86Sr croît au cours du temps par suite de
l’enrichissement en strontium 87 provenant de la désintégration du rubidium 87.
Les points représentatifs de la roche A se répartissent sur une droite de pente a. Si le granite B
s’était formé en même temps que le granite A, les points représentatifs des minéraux de ce
granite B se trouveraient sur une droite de même pente : ce n’est pas le cas.
En conséquence, les granites A et B ne se sont pas formés à la même époque et, la pente b
relative aux rapports isotopiques du granite B étant plus forte que la pente a, le granite B est
plus âgé que le granite A.
PARTIE 3 : Phénotype au niveau de l’organisme et hormones sexuelles (question 2B)
L’énoncé indique que la patiente, en dehors des organes génitaux externes et d’une partie du
vagin, la patiente ne possède pas d’organes génitaux féminins.
En revanche elle présente des testicules en position intra-abdominale mais qui ne produisent
pas de spermatozoïdes (pas de signes pubertaires) et ne possède pas de voies génitales mâles.
• Exploitation du document B
Le caryotype présente une paire de gonosomes XY : il s’agit donc d’un caryotype masculin. Il
y a donc contradiction entre le phénotype et le caryotype de la personne.
• Exploitation du document C
La patiente présente:
- une concentration de LH et de FSH élevée, supérieure à la concentration normale
maximale ;
- une concentration de testostérone élevée (supérieure à la normale maximale) prouvant la
capacité du testicule à répondre aux gonadostimulines FSH et LH.
Les concentrations élevées, à la fois de gonadostimulines et de testostérone, indiquent que le
rétrocontrôle négatif de la testostérone sur l’hypophyse ne fonctionne pas.

© Hatier, 2003 4
• Exploitation du document D :
Il fournit l’explication à l’ absence de réponse de l’hypophyse : la personne possède un gène
codant pour le récepteur à la testostérone qui présente une mutation, le 125e codon GAG
étant transformé en un codon TAG = STOP .
Par suite de la présence de ce codon STOP, le récepteur est raccourci et non fonctionnel. En
l’absence de récepteur fonctionnel, les cellules hypophysaires, cibles de la testostérone se
comportent comme en l’absence de celle-ci.
Bilan
Cette personne possède un caryotype masculin ; durant la vie fœtale la différenciation des
gonades s’est donc faite en testicules qui ont sécrété deux hormones : la testostérone et
l’AMH. Les récepteurs à la testostérone étant non fonctionnels, les canaux embryonnaires
masculins (canaux de Wolff) et les organes génitaux externes n’ont pas réagi à la
testostérone ; les canaux de Wolff ont dégénéré tandis que les organes génitaux externes se
féminisaient. On trouve là l’explication du phénotype féminin à la naissance.
En revanche, les voies féminines (canaux de Müller) ont répondu à la présence d’AMH et ont
disparu. d’où un phénotype féminin sans voies génitales féminines.
A la puberté, la commande hypothalamo-hypophysaire des testicules fonctionne bien ; ceux-ci
y répondent par une sécrétion de testostérone mais celle-ci reste inefficace par manque de
récepteurs fonctionnel sur les cellules cibles, d’où l’absence de puberté.
Il en résulte le phénotype observé : phénotype féminin sans voies génitales féminines et sans
signe pubertaire avec un caryotype XY.

© Hatier, 2003 5
SUJET 2
Partie 1 : Le brassage génétique
Introduction
Au sein des populations de chaque espèce, il y a un polymorphisme des gènes de sorte que les
individus diploïdes sont hétérozygotes pour environ 10% de leurs gènes. Jouant sur cette
hétérozygotie, les méioses qui président à la formation des gamètes haploïdes, assurent un brassage
génique tel que chaque gamète issu d’une méiose possède une association originale des allèles des
gènes de l’espèce.
Nous allons envisager un des mécanismes de ce brassage génique en considérant les méioses chez un
organisme hétérozygote pour trois gènes situés sur le même chromosome. Cela élimine l’intervention
du brassage interchromosomique ; seul le brassage intrachromosomique est donc envisagé.
I. La localisation chromosomique des allèles dans les cellules de l’organisme qui subissent la
méiose (figures 1)
Toutes les cellules de l’organisme et donc toutes les cellules mères de gamètes ont les mêmes
allèles des gènes. Les trois gènes envisagés situés sur le même chromosome sont donc liés. Plus
précisément, pour chaque gène, chaque cellule possède deux allèles, l’un sur un chromosome, l’autre
sur l’homologue, au même locus.
Considérons alors trois gènes A, B et D pour lesquels l’organisme est hétérozygote et désignons par
A et a ; B et b ; D et d les allèles de ces gènes.
Une disposition possible de ces allèles sur les chromosomes des cellules de l’organisme est celle
schématisée par la figure 1.
Figure 1. Disposition des allèles sur les chromosomes homologues.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
1
/
22
100%