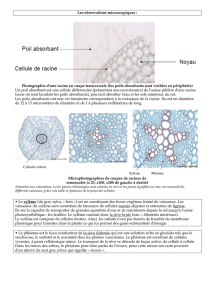
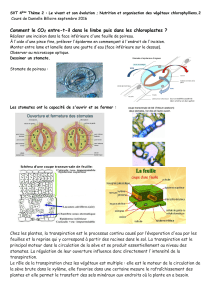
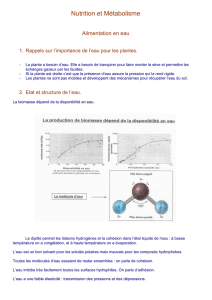
eau - Free

1
Schéma général :
fonctionnement intégré
de la plante
pour la circulation des sèves

2

3
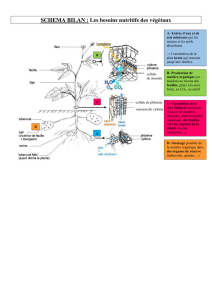
Le fonctionnement intégré
de la plante vasculaire
est illustré dans la figure suivante
par les échanges
entre les voies de transit
d’eau + sels minéraux (xylème)
et celles de transit des solutés minéraux
et organiques (phloème).

4
Mécanismes de transport chez les végétaux
Le transport des sèves ou des secrétions s'effectue à trois niveaux
chez les végétaux :
1- la captation de l'eau minéralisée et la sécrétion des solutés à
l'échelle cellulaire, telle l'absorption de l'eau et des minéraux du sol
par les poils absorbants des cellules d'une racine (transport au
niveau cellulaire);
2- le transport de nutriments d'une cellule à l'autre
dans un tissu ou organe, tel le transport de glucides
des cellules photosynthétiques d'une feuille
jusqu'aux tubes criblés du phloème (transport radial) ;
3- le transport des sèves dans le xylème et le phloème de la plante
entière (transport vertical).

5
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
1
/
67
100%