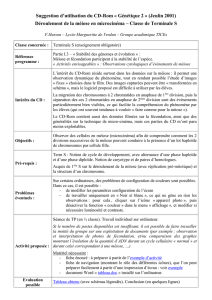
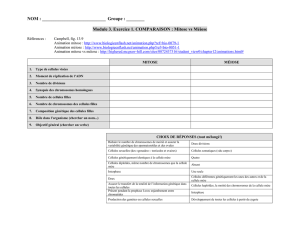
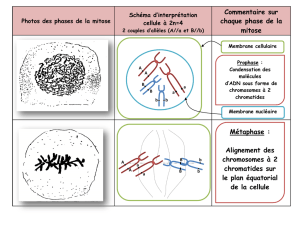
Chapitre 2: Méiose et fécondation : Stabilité et

Méiose et fécondation :
Stabilité et variabilité
Comment assurer le maintien
constant du caryotype au cours
des générations ?

I/Méiose et fécondation participent
à la stabilité de l’espèce.
A/ Le cycle biologique (=de
développement) des Mammifères.
Représenter le cycle de vie d'un Mammifère
tel que l'Homme, en précisant bien la
ploïdie (nombre n ou 2n de chromosome)
des cellules mises en jeu.

L’étude des caryotypes (docs 2 et 3p99)
permet de déterminer la ploïdie d’une
cellule.
•Les cellules somatiques possèdent deux
exemplaires de chromosomes : les
chromosomes identiques sont dits
homologues et sont organisés par paire ;
la cellule est diploïde.
•Les cellules sexuelles ne présentent qu’un
seul exemplaire de chaque chromosome :
la cellule est haploïde.

À compléter à l'aide du doc 1p98

Chez les mammifères, au cours du cycle de
développement, une phase diploïde (2n) alterne
avec une phase haploïde (n) ;la phase diploïde
est dominante.
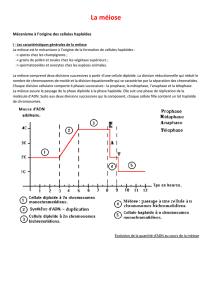
La méiose assure le passage de la phase diploïde
àla phase haploïde (2n →n) : elle assure la
formation des gamètes (ovules et
spermatozoïdes).
La fusion de ces gamètes, au cours de la
fécondation, est àl’origine d’une cellule
diploïde (n+n →2n) :la cellule-œuf.
Cette cellule va connaître des mitoses et sera à
l’origine d’un nouvel individu.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
1
/
38
100%