coursHPM-M2-RIA-nov05 - LIRIS

Réseaux de neurones temporels :
dynamique et apprentissage
Hélène Paugam-Moisy
Professeur à l’Université Lyon 2
Institut des Sciences Cognitives

Les réseaux de neurones naturels

Cerveau, neurones et intelligence
Le cerveau humain est constitué d’environ
1011 neurones ;
en moyenne, chaque neurone a de l’ordre de
104connexions.
Chez les animaux, la
place occupée par le
cortex semble être
proportionnelle au
développement des
facultés intelligentes.

Neurones biologiques / naturels
Les neurones forment des réseaux de communication complexes, chaque
neurone établissant de très nombreuses connexions avec d'autres.
vibrisses du rat : neurones sensitifs => thalamus
(White et Peters )neurones thalamiques => cortex

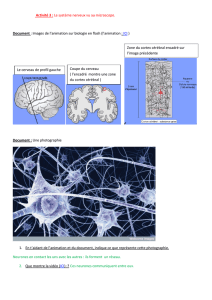
Neurones biologiques / naturels
gallerie de photos...
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
1
/
52
100%