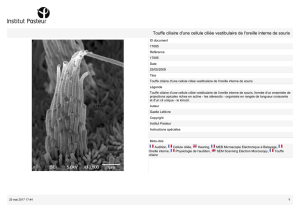
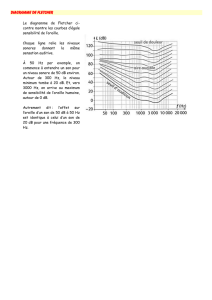
Mécano-sensibilité active des cellules ciliées de l`oreille interne



20Hz
20kHz
100kHz
4f
f0,5
20µP a 20P a
IdB = 20 log δP
δPref =
10 log δE
δEref Patm = 105P a δPref = 20µP a δEref = 10−12W/m2
0dB

=⇒=⇒
200nm

Vin −Vout =−60mV
Ca2+ K+
Ca2+
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%