université ouverte 2014-‐2015 - Laboratoire Matière et Systèmes

!"#$%&'"((")*"$(+)
,#-.(#/.'(")0#12(")"/)
3+4/25"4)6.578"9"4:)
;$'<"(4'/=)&#('4)>'?"(./)
@A7BCCDDDE54FEG$'<%7#('4%?'?"(./EH(C
I@"$(+C)
;JKLMN3KOP)Q;LMNOM)RSTU%RSTV)
Du#neurone#au#cerveau:#
Une#promenade#dans#la#complexité#
#du#système#nerveux))

W.$F1.$4)F.578"9"4B)
)6.X$'1.$:)"57#/@'")"/)=5.1.$4)
!"G?')TR)0#(4)

N=4G5=)?G)F.G(4)7(=F=?"$/)
• ,"4).(X#$"4)?"4)4"$4)'$H.(5"$/)8")F"(<"#G)4G()
8Y"$<'(.$$"5"$/Z)F"A")'$H.(5#1.$)?.'/)[/(")
"$("X'4/(=")4.G4)G$")H.(5")=8"F/('\G")
• ,"4).?"G(4)]5.8=FG8"4).?.(#$/"4^:)8"4)'5#X"4)
]7@./.$4^)4.$/)/(#$4H.(5="4)7#()8Y=7'/@=8'G5).8H#F1H)
"/)8#)(=1$")
• 6"A")/(#$4H.(5#1.$)"4/)"_"F/G=")7#()?"4)
(=F"7/"G(4)5=/#-./(.7"4)]`)@=8'F"4^)
• ,#)8'#'4.$)?")8Y.?.(#$/:)8Y#-4.(71.$)?G)7@./.$)
7(.?G'4"$/)G$)F@#$X"5"$/)?")F.$H.(5#1.$)\G')<#)
'$?G'(")G$")5.?'aF#1.$)?G)7./"$1"8)?G)$"G(.$")
4"$4.('"8)

N=4G5=)?G)F.G(4)7(=F=?"$/)
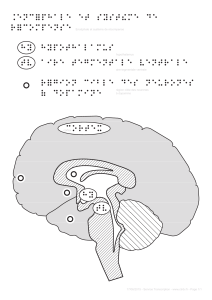
• bG))$'<"#G)?G)-G8-").8H#F1H)"/)?")8#)(=1$":)8"4)
$"G(.$"4)4"$4.('"84)4")7(.c"A"$/)4.G4)H.(5")?")
F#(/"4)
• ,Y'$H.(5#1.$)7#44")"$4G'/")?#$4)8"4)F.(/"9).8H#F1H)
]/"57.(#8)5=?'#$^)"/)<'4G"8)].FF'7'/#8^)
• >#$4)8"4)?"G9)F#4:)F"4)4/(GF/G("4)#$#8+4"$/)
8Y'$H.(5#1.$)?.$/)<#)[/(")"9/(#'/")8#)7"(F"71.$)
• >#$4)8")F.(/"9)<'4G"8:)F"4)'$H.(5#1.$4)F.$F"($"$/)
8"4)8'X$"4:)8"G()H(=\G"$F":)8"G().('"$/#1.$:)8#)
?.5'$#$F").FG8#'(":EE)

N=4G5=)?G)F.G(4)7(=F=?"$/)
• M$)#<#$/)?G)F.(/"9)<'4G"8)7('5#'(":)8Y'$H.(5#1.$)"4/)
"9/(#'/")#G)$'<"#G)?Y#'("4)47=F'#8'4="4)]/"57.(#8"4)
<"$/(#8"4^)
• Q$)?'41$XG")?"4)#'("4)?")("F.$$#'44#$F")?"4)
<'4#X"4:)?"4)"$<'(.$$"5"$/4:)?"4).-c"/4:)d)
• ,YKN0)H.$F1.$$"88")7"(5"/)8Y#$#8+4")?")F"4)=/#7"4:)
7"(5"A#$/)?")e)8'(")f)F")\G")<.'/)8")F"(<"#G)
• ,Y=8"F/(.7@+4'.8.X'")5.$/(")\GYG$)$"G(.$")7"G/)
(=#X'()g)G$")'5#X")F.578"9")
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
1
/
71
100%