Le tissu nerveux (deuxième partie)

Le tissu nerveux (deuxième partie) file:///C:/Documents%20and%20Settings/utilisateur/Bureau...
Le tissu nerveux (deuxième partie)
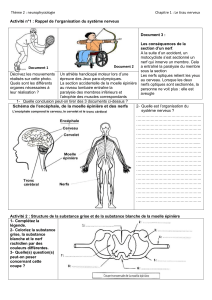
4.5. Architecture du système nerveux central
Le système nerveux central comprend l'encéphale enfermé dans la boîte crânienne et la moelle épinière qui occupe toute la longueur du canal
vertébral. Il est enveloppé par les méninges. Le système nerveux central est organisé en substance blanche et substance grise, disposées autour
d'un canal. La substance blanche est l'ensemble des axones myélinisés qui parcourent le système nerveux central dans toutes les directions et
sont associés en faisceaux. Ce sont les lipides de la myéline qui lui donnent son aspect blanchâtre. La vascularisation y est peu développée.
La substance grise, grisâtre dans un encéphale observé à frais, contient les péricaryons des neurones. La vascularisation y est très
développée.
La distribution des substances grise et blanche varie selon les régions du système nerveux central. Dans la moelle, la substance grise est uniquement
centrale; elle est entourée de substance blanche. Dans l'encéphale, au contraire, la substance grise est localisée dans des noyaux gris profonds et dans
le cortex périphérique du cervelet et du cerveau; la substance blanche y est sous-corticale.
4.5.1. Développement embryologique
Nous avons déjà vu que le système nerveux central provenait du tube neural. Celui-ci dérive d'une bande dorsale de l'ectoblaste qui se
déprime d'abord, sur toute la longueur de l'embryon, pour former la gouttière neurale. Les deux lèvres de la gouttière se rapprochent progressivement
pour délimiter un canal qui finalement se détache de l'ectoblaste.
Le tube neural s'allonge. Dans sa partie antérieure, qui est à l'origine de l'encéphale, il se dilate et forme trois vésicules: le prosencéphale, le
mésencéphale et le rhombencéphale. Le prosencéphale produira le télencéphale d'où naissent deux expansions latérales, à l'origine des hémisphères
cérébraux. La lumière du tube neural s'y dilatera en ventricules, cavités remplies de liquide céphalo-rachidien.
L'allongement du tube neural va de pair avec l'épaississement de sa paroi. Celle-ci, au départ, est composée de deux couches : la couche interne, ou
épendymaire, est épithéliale et limite la lumière; la couche externe est composée de cellules non différenciées. Certaines deviennent des cellules
gliales qui s'allongent et se disposent radiairement. Les autres, futurs neurones, se multiplient et migrent en périphérie pour former la substance grise.
Au cours de leur migration, elles glissent le long des prolongements des cellules gliales. Dans l'encéphale, celles qui migrent le plus loin de la couche
originelle constitueront les cortex; les vagues cellulaires successives sont à l'origine des couches cellulaires corticales superposées. D'autres migrent
sur de courtes distances et formeront les noyaux gris de la base.
Lorsqu'ils ont atteint leur position définitive, les neurones acquièrent un cône de croissance, qui s'allonge progressivement pour former l'axone. Les
axones de tous les neurones impliqués dans une fonction déterminée se développent en même temps de façon synchronisée. Ainsi s'explique la
formation des faisceaux qui constituent la substance blanche.
4.5.2. Types de neurones
Les neurones du système nerveux central ont les caractères généraux de tous les neurones; cependant, leur taille et leur forme varient en
fonction de leur localisation et de leur fonction.
Le neurone de Betz (1) est spécifique du cortex cérébral: son péricaryon est pyramidal, l'axone naît de sa base tandis que les dendrites émergent de
ses angles. Celle qui est issue de son sommet forme avec ses collatérales une sorte de panache qui atteint la surface corticale.
La cellule de Purkinje (2) est un neurone du cervelet dont le corps cellulaire a un peu la forme d'une poire. Les dendrites ont de très nombreuses
collatérales dont l'ensemble est disposé en espalier.
Certains neurones, appelés grains ou cellules granulaires (3), sont petits; leur noyau volumineux occupe presque tout le cytoplasme et leurs dendrites
sont courtes.
D'autres, les cellules de Cajal (4), sont fusiformes, leurs neurites étant parallèles à la surface du cortex.
4.5.3. Cellules gliales

Le tissu nerveux (deuxième partie) file:///C:/Documents%20and%20Settings/utilisateur/Bureau...
Les cellules gliales du système nerveux central sont les astrocytes, les oligodendrocytes, les cellules épendymaires et les
cellules de la microglie.
Les astrocytes sont les cellules de soutien du système nerveux central. Le corps cellulaire et ses nombreux prolongements sont hérissés de fines
expansions qui lui donnent l'aspect d'une pelote d'épingles. Ils ne peuvent être mis en évidence que par des techniques appropriées. Ici, dans la
substance blanche, les prolongements de l'astrocyte sont longs et grêles; c'est pourquoi il est dit fibreux. Dans la substance grise, les prolongements
de l'astrocyte sont courts et épais : il est dit protoplasmique.
Les astrocytes interviennent dans les échanges entre les neurones et les vaisseaux. Les prolongements astrocytaires s'appliquent contre la surface des
capillaires. Il forment ainsi un manchon, la "membrane limitante gliale périvasculaire". Dans cette micrographie électronique, les prolongements des
astrocytes (A) sont des plages claires appliquées contre la paroi d'un capillaire cérébral (C). Ils n'en sont séparés que par la membrane basale du
capillaire et le revêtement externe de l'astrocyte, qui sont désignés par des flèches.
D'autres prolongements s'accolent aux péricaryons et aux neurites et s'insinuent même près des fentes synaptiques. Chaque astrocyte (A) forme ainsi
avec un neurone (N) et un capillaire (C) une unité angio-glio-neurale. Elle fut pendant longtemps considérée comme le support morphologique de la
barrière hémato-encéphalique, sorte de filtre qui empêche certaines molécules circulant dans le sang d'atteindre les espaces intercellulaires du
système nerveux central. On sait aujourd'hui que ce sont les cellules endothéliales elles-mêmes qui sont responsables de cette perméabilité sélective.
L'oligodendrocyte est une petite cellule, à noyau dense et à prolongements courts. Il est responsable de la myélinisation et donc l'équivalent de la
cellule de Schwann dans le système nerveux périphérique. Voici un oligodendrocyte en microscopie électronique. Son noyau est dense. Chaque
prolongement est en rapport avec un neurite et lui fournit une gaine de myéline. Il existe des différences entre la cellule de Schwann du système
nerveux périphérique et l'oligodendrocyte du système nerveux central. Dans la cellule de Schwann, un neurite unique est inclus dans la cellule qui lui
fournit sa gaine de myéline. Par contre, un seul oligodendrocyte assure la myélinisation de plusieurs neurites en émettant des prolongements en
forme de voiles dont le bord s'invagine pour envelopper un axone.
Voici un axone myélinisé du système nerveux central. La périodicité de la gaine de myéline est identique à celle décrite dans le système nerveux
périphérique. Des incisures de Schmidt-Lantermann interrompent la striation périodique.
Les cellules épendymaires limitent la lumière du tube neural ou canal épendymaire. Dans la moelle, ce canal est collabé, les cellules sont
atrophiées et leur pôle basal est irrégulier. Dans l'encéphale, il est délimité par un épithélium cubique simple; dans les quatre ventricules, les cellules
épendymaires sont ciliées.
Dans le toit des ventricules (V), l'épithélium forme les plexus choroïdes, séries de replis richement vascularisés qui se projettent dans la cavité. Leurs
cellules sont glandulaires, elles ont de longues microvillosités et leur cytoplasme est riche en mitochondries. Elles produisent le liquide
céphalo-rachidien qui coule dans les ventricules et dans les espaces sous-arachnoïdiens des méninges. Ce liquide contient quelques protéines, des
sels minéraux et du glucose. Son rôle est surtout mécanique : il allège le cerveau qui flotte littéralement dans la boîte crânienne et le protège des
traumatismes.
Les cellules de la microglie sont très ramifiées, elles possèdent un noyau allongé, dense, de petite taille et des prolongements fins et très ramifiés.
Elles sont mobiles et leur pouvoir phagocytaire est important.
4.5.4. Méninges
Les méninges sont des lames tissulaires qui enveloppent le système nerveux central. Il en existe deux : l'interne, ou leptoméninge,
est d'origine ectoblastique; l'externe, ou pachyméninge, est de nature conjonctive et d'origine mésoblastique. Leur composition est la même dans le
crâne, autour de l'encéphale, et dans le canal rachidien, autour de la moelle épinière.
La leptoméninge comprend la pie-mère et l'arachnoïde. La pie-mère (P), composée de cellules aplaties, adhère intimement au système nerveux
central dont elle épouse tous les replis. L'arachnoïde (A), est un tissu aréolaire dont les travées, tapissées par un épithélium pavimenteux simple,
contiennent des fibres collagènes, des cellules conjonctives et des capillaires. Ses espaces sont remplis de liquide céphalo-rachidien.
La pachyméninge, ou dure-mère, est un tissu conjonctif dense. Elle contient les grandes veines du crâne, les sinus veineux (S). Elle est séparée de
l'arachnoïde par un espace virtuel, l'espace sous-dural. Dans le crâne, elle se confond avec le périoste et est traversée par des expansions
arachnoïdiennes (A) qui entrent en relation étroite avec l'endothélium des vaisseaux et assurent la résorption du liquide céphalo-rachidien. Dans le
canal vertébral, qui n'est pas représenté dans ce schéma, elle est séparée du périoste par l'espace épidural, rempli de tissu graisseux très vascularisé.
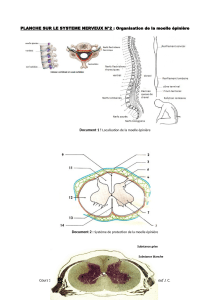
4.5.5. Moelle épinière
Dans la moelle épinière, la substance blanche, colorée en brun foncé par une imprégnation à l'argent, est périphérique et formée de
faisceaux d'axones myélinisés dont l'épaisseur augmente au fur et à mesure que l'on se rapproche de la région crâniale. Elle est subdivisée
topographiquement, dans chaque moitié de la moelle, en trois cordons : antérieur, latéral et postérieur. Une commissure unit les deux cordons
antérieurs. Cette subdivision fasciculaire n'est pas perceptible dans cette préparation.

Le tissu nerveux (deuxième partie) file:///C:/Documents%20and%20Settings/utilisateur/Bureau...
La substance grise est centrale. Elle est moins colorée et contient les corps cellulaires et des neurites non myélinisés.
La substance grise, en coupe transversale dans ce schéma, a la forme d'un H majuscule dont les branches sont appelées cornes. La partie centrale est
traversée par le canal épendymaire et s'appelle commissure grise. Les cellules multipolaires de la corne antérieure (c) sont des motoneurones. Leurs
axones quittent la moelle par la racine ventrale (e) pour se rendre dans le nerf rachidien (h). Ils innervent les fibres musculaires. Ce trajet est indiqué
en rouge dans le schéma. Leurs dendrites sont courtes et en relation avec des neurones-relais de la corne dorsale ou avec des neurones d'association.
Les cellules étoilées ou fusiformes de la corne dorsale (a) ont une signification sensitive. Le trajet à signification sensitive est indiqué en bleu dans le
schéma. Les stimuli périphériques sont recueillis par les dendrites des neurones dont le corps cellulaire est situé dans le ganglion rachidien (f). Les
axones de ces neurones ganglionnaires rejoignent la moelle par la racine dorsale (d) et font synapse avec les dendrites des neurones de la corne
dorsale. Les axones de ces neurones se dirigeront vers des centres supérieurs. Il existe, entre la corne antérieure et la corne postérieure, une petite
corne appelée médio-latérale (b). Elle contient de petites cellules à signification autonomique, c'est-à-dire indépendante des voies conscientes. Leur
axone quitte la moelle par la racine antérieure puis rejoint un ganglion prévertébral (g) par le rameau communicant blanc (i). Dans ce ganglion
autonomique, l'axone s'articule avec les dendrites d'un ou de plusieurs neurones dont les axones sont les voies autonomiques efférentes. Certains
regagnent le nerf rachidien par le rameau communicant gris (j). D'autres entrent dans la constitution des nerfs viscéraux; d'autres unissent les
différents ganglions de la chaîne prévertébrale. Leur trajet est représenté en noir dans le schéma. Enfin, il existe, répartis dans toute la substance
grise, des neurones appelés intercalaires qui unissent différents segments de la moelle.
4.5.6. Cervelet
Le cervelet contrôle l'équilibre et coordonne le tonus postural et les mouvements volontaires ou involontaires. Il comprend le vermis
médian et deux hémisphères latéraux. Il est relié à la face dorsale du tronc cérébral par trois paires de pédoncules. La substance blanche y forme un
centre médullaire d'où partent des lames médullaires qui se ramifient en un grand nombre de lamelles périphériques. La substance grise comprend
quatre paires de noyaux profonds et un cortex périphérique qui forme de nombreux replis.
Voici quelques replis du cortex cérébelleux. Chacun contourne une lamelle de substance blanche où les fibres nerveuses ont été colorées en noir par
une imprégnation argentique. Même à ce grossissement relativement faible, il est possible de reconnaître les trois couches du cortex cérébelleux. La
couche la plus externe ou moléculaire contient peu de cellules. La couche moyenne ou ganglionnaire est formée d'une rangée de volumineux corps
cellulaires. La couche interne ou granuleuse est la plus épaisse et la plus colorée. Elle contient de très nombreux grains.
Un plus fort grossissement permet d'observer le détail des trois couches du cortex cérébelleux. Les cellules volumineuses, au centre de l'image, sont
les cellules de Purkinje, hautes d'environ 60 µm, piriformes et disposées en une rangée régulière. Leur volumineux noyau est sphérique.
La schématisation du cortex cérébelleux permet d'expliquer la signification de chaque couche.
La couche moléculaire (A) est avant tout un enchevêtrement de fibres nerveuses. Elle contient de petits neurones multipolaires ou cellules étoilées
dont les courts neurites sont tangentiels à la surface. Les plus profonds ont un long axone, parallèle à la surface, dont les collatérales dessinent une
corbeille synaptique autour du corps des cellules de la couche sous-jacente.
La couche ganglionnaire (B) est formée par l'alignement des cellules de Purkinje. Leur dendrite s'enfonce dans la couche moléculaire et atteint la
superficie du cortex. Elle se ramifie en espalier et occupe en surface une petite zone rectangulaire. L'axone s'enfonce dans la substance blanche et se
dirige vers les noyaux gris profonds.
L'axone des grains de la couche granuleuse (C) remonte vers la surface et fait synapse avec les dendrites des cellules étoilées et des cellules de
Purkinje. Leurs dendrites s'articulent avec les fibres en provenance de la moelle et du bulbe. La couche granulaire contient en plus des neurones qui
associent différentes zones du cortex cérébelleux.
4.5.7. Cortex cérébral
Dans les hémisphères cérébraux, la substance grise comprend les noyaux profonds et le cortex cérébral dont la surface, grâce aux
très nombreux replis, représente près de 2,2 m2. Il contient environ dix milliards de neurones. Les plis profonds délimitent les lobes; d'autres, moins
profonds, divisent les lobes en circonvolutions. La substance blanche du cerveau est sous-corticale; elle est formée d'un lacis d'axones myélinisés
dont les faisceaux se ramifient en périphérie. Le trajet de ces faisceaux dépend des relations qu'ils établissent entre les différentes parties du système
nerveux. Les fibres associatives relient deux zones corticales d'un même hémisphère. Les fibres commissurales relient des zones corticales des deux
hémisphères et les fibres de projection, afférentes ou efférentes, connectent le cortex avec des centres sous-corticaux.
La structure histologique du cortex cérébral est complexe. On peut la décrire à partir de coupes histologiques colorées selon des techniques qui
mettent les corps cellulaires en évidence. De ce point de vue, dit cytoarchitectonique, on peut, déjà à faible grossissement, identifier plusieurs
couches cellulaires.
En colorant la myéline, on met en évidence des couches de fibres successives, parallèles à la surface. De ce point vue, dit myéloarchitectonique, on
distingue dans le cortex cérébral plusieurs stries. On colore également les fibres radiées myélinisées, perpendiculaires au cortex, qui y pénètrent ou
qui en sortent.
Du point de vue de la cytoarchitectonie, on distingue dans le cortex cérébral six couches qui sont, de dehors en dedans et donc de haut en bas dans le
schéma :
la couche moléculaire qui ne contient que quelques cellules de Cajal,1.

Le tissu nerveux (deuxième partie) file:///C:/Documents%20and%20Settings/utilisateur/Bureau...
la couche granulaire externe composée de grains serrés,2.
la couche des cellules pyramidales petites et moyennes,3.
la couche granulaire interne composée de grains serrés,4.
la couche ganglionnaire qui contient les grandes cellules pyramidales de Betz,5.
la couche des cellules polymorphes au grand axe perpendiculaire à la surface corticale.6.
Du point de vue de la myéloarchitectonie, on distingue, de dehors en dedans :
le plexus d'Exner qui occupe la couche moléculaire,1.
la strie de Bechterew située entre la deuxième et la troisième couche cellulaire,2.
la strie externe de Baillarger qui occupe la couche granulaire interne,3.
la strie interne de Baillarger située dans la partie profonde de la cinquième couche.4.
Les fibres radiées, perpendiculaires à la surface du cortex, unissent ces différentes zones.
Ce schéma explique les rapports de ces différentes zones. Au centre, une zone corticale reliée, à gauche, avec une zone corticale voisine du même
hémisphère et, à droite, avec une zone corticale de l'hémisphère opposé. En noir, les cellules réceptrices, en rouge, les cellules effectrices.
Les couches 1, 2 et 4 sont réceptrices. Les cellules de Cajal, dans la couche 1 moléculaire, associent les axones des grains de la couche 2 et les
dendrites des cellules pyramidales des couches 3 ou 5. Les grains de la couche 2 reçoivent des informations d'autres zones corticales du même
hémisphère. Leurs axones sont en relation avec les neurones de la couche 1 ou directement avec les cellules effectrices. Les grains de la couche 4
granulaire interne reçoivent des informations de zones sous-corticales. Leurs axones se dirigent vers les couches effectrices 3 et 5.
Les couches 3, 5 et 6 sont effectrices. La couche 3 des cellules pyramidales petites et moyennes émet des fibres associatives destinées aux couches 2
et 5 d'autres zones du même hémisphère. Les grosses cellules pyramidales de la couche ganglionnaire 5 émettent des axones destinés aux centres
sous-corticaux. C'est donc la couche la plus développée dans les aires corticales motrices. Les cellules polymorphes de la couche 6 envoient leurs
axones, fibres commissurales, vers la zone 5 de la région homologue de l'hémisphère opposé.
suite du TP...
1
/
4
100%