VISION - - Neurophysiologie visuelle », Encyclopædia Universalis

© Encyclopædia Universalis France
VISION - Neurophysiologie visuelle
Article écrit par Jean BULLIER
Prise de vue
La vision est certainement la modalité sensorielle dominante chez l'homme. La compréhension des bases
physiologiques de cette fonction a donné lieu à de nombreuses études réalisées principalement chez les vertébrés
supérieurs et plus particulièrement chez le singe, dont l'organisation du système visuel se rapproche le plus de celle de
l'homme. Cet article traite de la physiologie du traitement de l'information visuelle dans la rétine et dans les structures
sous-corticales et corticales. Les phénomènes de transformation du signal lumineux en signal électrique sont abordés dans
l'article VISION - Photoréception.
I-Rôle de la rétine
La rétine est la structure du système visuel pour laquelle la connaissance des corrélations entre structure et fonction est
la plus avancée. Comme nous le verrons, on arrive à expliquer les réponses de nombreux neurones rétiniens en fonction de
leurs connexions avec les autres éléments du réseau. La rétine des primates est constituée d'une série de couches
comportant différents types de neurones et leurs synapses, depuis les photorécepteurs qui constituent l'étage d'entrée du
signal jusqu'à l'étage de sortie formé par les cellules ganglionnaires dont les axones se projettent via le nerf optique vers les
centres supérieurs de traitement.
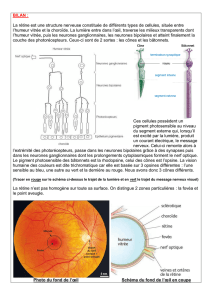
Les segments externes des photorécepteurs (cônes ou bâtonnets) sont logés dans l'épithélium pigmentaire, dont le rôle
est de faciliter la régénération des pigments photosensibles. Cet épithélium pigmentaire qui tapisse le fond de l'œil (fig. 1)
est entouré par la choroïde, couche riche en vaisseaux sanguins, et par la sclérotique fibreuse qui constitue l'enveloppe de
l'œil.

© Encyclopædia Universalis France
Rétine (vue en coupe)
Schéma d'une coupe transversale de rétine de primate
(épaisseur 0,5 mm environ). Les différents types de
neurones (cônes, bâtonnets, cellules bipolaires,
horizontales, amacrines, ganglionnaires) sont répartis dans
des couches cellulaires et synaptiques. Le haut de la figure
correspond à la partie externe de la rétine (proche de la
sclérotique qui enveloppe le globe oculaire). Les
…(Encyclopædia Universalis France)
Les cellules ganglionnaires étant situées au niveau de la partie interne de la rétine, proche du corps vitré qui remplit la
chambre postérieure du globe oculaire, les rayons lumineux doivent donc traverser toute l'épaisseur de la rétine pour
atteindre les photorécepteurs.
Rétine centrale et périphérique
Bien que tapissant de façon homogène la face interne du globe oculaire, la rétine est loin d'être une structure
homogène. Chez toutes les espèces animales, on trouve une région de plus forte densité de cellules photosensibles et de
neurones qui correspond à la zone de plus forte acuité visuelle. Chez l'homme et le singe, cette spécialisation est
particulièrement marquée : il existe une zone pour laquelle l'ensemble des neurones autres que les photorécepteurs sont
repoussés vers la périphérie, créant ainsi dans la rétine une sorte de petite indentation appelée fovéa (fig. 2). Dans la partie
centrale de la fovéa (sur un diamètre de 0,4 mm correspondant à 1,4 degré d'angle visuel), les vaisseaux sanguins sont
absents de la surface de la rétine, permettant ainsi un accès direct de la lumière aux photorécepteurs qui sont
exclusivement des cônes de type L et M (voir définition ci-après) atteignant des densités très importantes (fig. 3). Au-delà de
cette partie centrale de la fovéa, on trouve un mélange de cônes de type M, L et S (les cônes S sont 20 fois moins
nombreux que l'ensemble des M et L dans la rétine, alors que le rapport L/M varie de 1 à 4 suivant les individus). Enfin,
au-delà de la fovéa, la densité de bâtonnets augmente avec l'excentricité pour atteindre son maximum vers 15 degrés
d'excentricité (fig. 3), si bien que, dans la rétine périphérique, la densité des bâtonnets est beaucoup plus élevée que celle
des cônes. Cette très forte hétérogénéité de la rétine a des conséquences fonctionnelles importantes : la vision de forte
acuité est limitée à quelques degrés au centre du champ visuel, la perception des couleurs est très mauvaise dans la partie
du champ visuel correspondant à la rétine périphérique et la zone de meilleure sensibilité à des faibles niveaux de
luminance est située à environ 15 degrés d'excentricité. Cette hétérogénéité va se retrouver dans toutes les structures de
traitement de l'information visuelle avec, en particulier, une très forte quantité de neurones traitant les 5 degrés centraux du
champ visuel.

© Encyclopædia Universalis France
Globe oculaire
Coupe horizontale dans un globe oculaire droit humain. La
papille optique ou tache aveugle correspond à la région où
se rassemblent les axones des cellules ganglionnaires et qui
ne comporte donc aucun photorécepteur.(Encyclopædia
Universalis France)
Photorécepteurs rétiniens
Courbe de densité des cônes et bâtonnets en fonction de la
distance angulaire par rapport au centre de la fovéa. Noter
l'absence de photorécepteurs au niveau de la tache aveugle
(modifiée à partir de Rodieck 1998).(Encyclopædia
Universalis France)
Cônes et bâtonnets
Les deux types de photorécepteurs, cônes et bâtonnets, qui existent dans la rétine des vertébrés ont une rôle différent
(fig. 3). Les bâtonnets possèdent une excellente sensibilité à des bas niveaux de luminance (on admet qu'un bâtonnet peut
être excité par un seul photon), mais donnent des réponses qui saturent au-delà d'un niveau d'éclairement de quelques
candelas par mètre carré (cas d'une pièce faiblement éclairée). Ils sont donc de peu d'utilité pour la vision diurne qui est
assurée par le fonctionnement des cônes, lesquels ne saturent pas et peuvent coder des niveaux de luminance allant
jusqu'à plusieurs milliers de candelas par mètre carré (correspondant à la luminance des objets à l'extérieur par une journée
ensoleillée).
Les cônes diffèrent par leur courbe de sensibilité spectrale (cf. VISION - Photoréception). Les vertébrés possèdent
généralement deux types de cônes : le cône S, dont la courbe de sensibilité spectrale a un pic situé vers 450 nanomètres, et
le cône M, dont le pic de sensibilité se situe vers 550 nanomètres. Ces deux types de cônes permettent une capacité limitée
de vision des couleurs chez la plupart des espèces de vertébrés. Chez certains poissons, oiseaux et primates diurnes, on
trouve un plus grand nombre de cônes permettant une perception plus fine des différences de couleur entre les objets. Chez
les primates, on distingue trois types de cônes, S, M et L. Le cône L diffère très peu du cône M en termes de structure
moléculaire et de position du pic de sensibilité, ce qui est important pour limiter les effets des aberrations chromatiques de
l'image rétinienne (les pics de sensibilité des cônes S, M et L chez l'homme sont à 420, 530 et 565 nm et à 500 nm pour les
bâtonnets).

© Encyclopædia Universalis France
Types de cellules ganglionnaires
En étudiant la morphologie et la fonction des cellules ganglionnaires de la rétine, on peut distinguer trois types de
neurones, les cellules alpha, les cellules bêta et les cellules gamma (Watanabe et Rodieck, 1989 ; Wassle et Boycott, 1991).
Les cellules bêta (également appelées cellules naines ou midget en anglais) ont un corps cellulaire et un champ dendritique
de petite taille et sont activées par un cône (dans la partie fovéale du champ visuel) ou plusieurs. Ce sont les cellules
responsables de la vision des détails et elles participent également à la vision des couleurs. Les cellules alpha (également
appelées parasol du fait de la forme de leur arborisation dendritique) représentent approximativement 10 p. 100 des cellules
ganglionnaires chez le singe, ont un soma et un champ dendritique de plus grande taille que les cellules bêta, sont
importantes pour la vision de la forme et du mouvement, mais ne participent pas à la vision des couleurs. Enfin, les cellules
gamma, qui constituent également 10 p. 100 du total des cellules ganglionnaires chez les primates, ont des morphologies
variées et des propriétés mal connues pour la plupart.
Le champ récepteur
Prenons l'exemple d'une cellule ganglionnaire de type alpha ou bêta. On caractérise sa réponse à la stimulation
photique en éclairant par de petites taches de lumière la rétine ou un écran placé dans le champ visuel. Comme toutes les
cellules ganglionnaires, alpha ou bêta répond au changement de luminance (augmentation ou diminution) et relativement
peu à la présentation continue de la lumière. Cela est lié au fait que, au cours de l'exploration visuelle, l'œil se déplace
constamment d'un point à un autre et que, même lorsque le sujet fixe un point, l'œil est sujet à des micromouvements qui
font se déplacer sur la rétine l'image d'une tache lumineuse fixe dans le champ visuel. On caractérise donc les aspects
fonctionnels d'un neurone en déterminant les caractéristiques spatiales et temporelles de la réponse de ce neurone à des
stimuli dont la luminance ou la couleur varient dans le temps de façon sinusoïdale ou carrée.
Dans le domaine spatial, on définit le champ récepteur qui est une notion fondamentale pour l'ensemble de la recherche
en neurophysiologie de la vision. Le champ récepteur est la région du champ visuel dans lequel une stimulation lumineuse
transitoire permet de modifier la fréquence de décharge des potentiels d'action du neurone étudié. On observe deux types
de cellules ganglionnaires alpha ou bêta : celles qui donnent une réponse de type on et celles qui donnent une réponse de
type off. La réponse de type on correspond à une augmentation de la fréquence de décharge du neurone lorsque la
luminance d'une petite tache de lumière est augmentée dans une petite région du champ visuel (de l'ordre de quelques
dixièmes de degré d'angle visuel) appelée centre du champ récepteur (fig. 4, stimulus 2). La réponse de type off est une
augmentation de la fréquence de décharge lorsque la luminance est diminuée dans le centre du champ récepteur (fig. 4,
stimulus 2).

© Encyclopædia Universalis France
Cellules ganglionnaires rétiniennes : réponses de
type ON et OFF
En a, réponses d'une cellule ganglionnaire de type ON à
différents stimuli activant le centre du champ récepteur
(stimuli 2 et 4). La réponse de ce neurone de type ON à une
diminution de luminance dans le centre du champ récepteur
est une diminution de la fréquence de décharge des
potentiels d'action (stimulus 4). En …(Encyclopædia
Universalis France)
Si maintenant on agrandit progressivement la tache de lumière qui sert à stimuler la cellule ganglionnaire, on s'aperçoit
que la réponse augmente jusqu'à ce que la tache de lumière occupe la totalité du centre du champ récepteur, mais qu'elle
diminue lorsqu'on accroît la taille du stimulus au-delà de cette limite. Cette diminution de la réponse est due au fait qu'il
existe une région appelée pourtour du champ récepteur dont la stimulation conduit à diminuer la réponse du neurone. Cet
effet inhibiteur caractérise l'interaction antagoniste entre centre et pourtour (fig. 4). La conséquence fonctionnelle de cet
antagonisme est qu'une cellule ganglionnaire répond très peu à la stimulation par une tache lumineuse qui couvre le centre
et le pourtour (fig. 4, stimulus 6). En d'autres termes, la cellule ganglionnaire code le contraste de luminance (et de couleur
dans le cas des cellules bêta de la fovéa) entre le centre et le pourtour.
Réponses on et off et interactions entre centre et pourtour
Le type de réponse on ou off et les interactions entre centre et pourtour des cellules ganglionnaires sont le résultat de
spécialisations fonctionnelles tout au long de la chaîne de traitement dans le réseau rétinien (fig. 5). Contrairement aux
cellules ganglionnaires qui émettent des potentiels d'action, les autres neurones rétiniens ne répondent aux stimuli visuels
que par des augmentations (dépolarisation) ou des diminutions (hyperpolarisation) de leur potentiel de membrane (l'intérieur
d'un neurone a un potentiel de l'ordre de — 60 mV par rapport à l'extérieur). Dans un photorécepteur de vertébré maintenu
dans l'obscurité, il existe un courant continu, dû à des entrées d'ions sodium, qui contrôle le potentiel de membrane et le
maintient à un niveau constant (typiquement — 40 mV). L'énergie lumineuse transmise au segment externe diminue ce
courant, ce qui se traduit par une hyperpolarisation de la membrane (à — 70 mV par exemple ; cf. VISION -
Photoréception).
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%