LIRE - UNamur

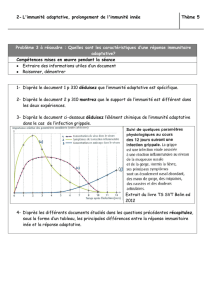
Les prix Nobel
de Physiologie – Médecine 2011
La mouche, la souris et la cellule dendritique
M T, J-J L
matthieu.terwagne@fundp.ac.be | jean-jacques.letesson@fundp.ac.be
1. Avant propos
«So, Naturalists observe, a ea
Hath smaller eas that on him prey;
And these have smaller still to bite ’em;
And so proceed ad innitum»
J S
On POETRY: A RHAPSODY ()
1.1 De Jonathan Swift…
Jonathan Swift avait raison quand il écrivait ces vers. La vie est relation
trophique et symbiose et ce, depuis la nuit des temps. Le terme symbiose est
assez vaguement déni et, pour cette raison, il a été appliqué à un large éven-
tail de taxons végétaux, fungiques, animaux et bactériens. On peut même
armer, sans crainte de se tromper, que tous les êtres vivants servent d’hôtes
. Le terme symbiose vient du Grec συμβʹiωσiς, (“vivre ensemble”) et, au sens large, fait
référence à la relation entretenue par un organisme d’une espèce donnée avec un orga-
nisme d’une espèce diérente, appelée “hôte”. Cette relation peut être bénéque (on
parle alors de mutualisme), neutre (commensalisme) ou détrimentale (parasitisme) pour
l’hôte.
Revue des Questions Scientiques, , (-) : -

à au moins un symbionte. Cette perception du vivantcomme “avant tout
symbiotique”est en plein essor et risque bien de bouleverser complètement la
perception que les biologistes ont du monde ainsi que l’interprétation qu’ils
font du vivant.
Une des implications sous-jacentes à toute relation symbiotique est qu’elle
nécessite une “communication structurée”, un échange bilatéral d’informa-
tions réglant l’établissement et le maintien de cet équilibre relationnel entre
espèces vivantes. Que l’échelle soit moléculaire ou macroscopique, que le si-
gnal soit physique ou chimique, la communication nécessite émission, récep-
tionet réaction adaptée au stimulus.
1.2 …à nos jours
Comme il semble bien que ce type de relation symbiotique soit ancestral,
alors, évolutivement parlant, tout être vivant doit contenir les informations
génétiques nécessaires à la mise en place des mécanismes de perception et de
réaction aux signaux de “communication” qu’implique cette symbiose. Si
l’on se focalise sur les relations (parasitaires ou non) qu’entretiennent les êtres
vivants multicellulaires avec les microorganismes (bactéries, virus ou proto-
zoaires), alors les plantes et les animaux doivent posséder des récepteurs ca-
pables de percevoir la présence de ces microorganismes an d’élaborer une
réponse que l’on qualie d’immune. Cette réaction vise à préserver l’intégrité
de l’hôte, non seulement en éliminant les pathogènes, mais également en
maintenant l’homéostasie entre l’hôte et sa microore commensale, égale-
ment appelée microbiote.
. (a) Price, P.W. . Evolutionary Biology of Parasites. Princeton University Press, Prin-
ceton. (b) Windsor, D. A. . Most of the species on Earth are parasites. Interna-
tional Journal for Parasitology (): –. (c) Poulin, R. . Evolutionary
Ecology of Parasites: second edition. Princeton University Press, Princeton.
. (a) McFall-Ngai, M. . Are biologists in “future shock?” Symbiosis integrates biol-
ogy across domains. Nat. Rev. Microbiol. :–. (b) Rohwer, F., and M. Youle.
. Consider something viral in your research. Nat. Rev. Microbiol. :–.
. Cette idée se retrouve dans la formule prémonitoire de Max Delbrück, en : “Any
living cell carries with it the experience of a billion years of experimentation by its ancestors”.
. Medzhitov, R. . Recognition of microorganisms and activation of the immune
response. Nature : -.
. Littman, D.R. and Pamer, E.G. . Role of the commensal microbiota in normal and
pathogenic host immune responses. Cell Host Microbe : -.

: , …
C’est de cela dont il est question dans l’attribution du prix Nobel de
Physiologie ou Médecine de : la découverte de récepteurs et de méca-
nismes susceptibles de détecter la présence (et la nature) de microorganismes
au sein de l’hôte, d’initier et d’orienter la réponse immune de celui-ci.
1.3 L’intenable prépondérance de l’immunité spécique
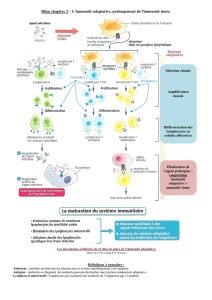
Le système immunitaire des vertébrés a depuis longtemps été décrit, de
manière un peu dogmatique, comme étant composé de deux branches: l’im-
munité innée, évolutivement ancienne et procurant une défense immédiate,
et l’immunité adaptative, également qualiée de hautement spécique, qui
procure une réponse plus tardive mais aussi une mémoire immunologique.
Il y a encore quelques années seulement, les manuels d’immunologie ne
consacraient que quelques pages à l’immunité innée, et ce dans des termes
génériques tels que “non spécique” ou “naturelle” en décrivant brièvement
les barrières physico-chimiques, le système du complément et les phagocytes.
Venaient ensuite plusieurs chapitres décrivant en détail les lymphocytes T et
B de l’immunité adaptative, leur sélection clonale, tout en insistant fortement
sur la spécicité de leur réponse ainsi que sur le développement de lympho-
cytes mémoires qui permettront une réaction plus rapide, plus vigoureuse et
plus spécique lors d’un deuxième contact avec le même agent infectieux.
Notons qu’en plus de ce déséquilibre entre l’importance accordée par les ma-
nuels d’immunologie à l’immunité innée et l’immunité adaptative, les deux
sujets étaient traités indépendamment l’un de l’autre.
Force est de constater que le nombre de chapitres consacrés à chacune des
deux parties du système immunitaire était inversement proportionnel à leur
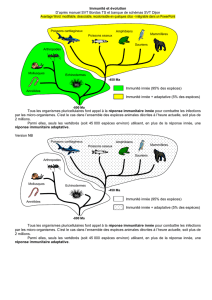
importance relative dans la survie des organismes. Au cours de l’évolution,
l’immunité adaptative n’est apparue qu’à partir des vertébrés à mâchoires
(gnathostomes). Elle ne concerne dès lors que quelques dizaines de milliers
d’espèces. À l’inverse, les invertébrés (dont certains fossiles vivants comme le
crabe fer à cheval Limulus polyphemus) et les plantes, qui constituent la majo-
rité des espèces vivantes sur Terre, comptent exclusivement sur leur système
. Il faut toutefois noter qu’une forme «non-classique» d’immunité adaptative a récem-
ment été décrite chez les agnathes dont les lamproies et les myxines sont les seuls repré-
sentants actuels (Herrin, B. R., and Cooper, M. D. (). Alternative adaptive immu-
nity in jawless vertebrates. J. Immunol. , -).

immunitaire inné pour assurer leur défense contre les agents pathogènes. De
plus, alors qu’il n’y a pas d’immunité adaptative sans immunité innée, l’in-
verse s’avère vrai. Ce seul fait trahi l’importance vitale des mécanismes innés
du système immunitaire.
1.4 Le juste retour de l’immunité innée sur le devant de la scène
Malgré son succès à l’échelle de l’évolution, l’immunité innée a donc
longtemps été traitée avec condescendance par les immunologistes, étant
considérée comme une simple mesure provisoire de défense en attendant que
l’immunité adaptative prenne le relais. Cette hégémonie du système immuni-
taire adaptatif a plus que probablement retardé, voire empêché la reconnais-
sance de l’importance de l’immunité innée. Et ce déséquilibre dans la
perception du système immunitaire s’est également traduit dans la répartition
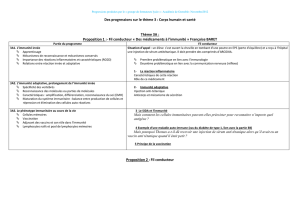
des prix Nobel de Physiologie ou Médecine décernés antérieurement à des
immunologistes (Table ). En eet, à l’exception des deux premiers Nobel
attribués à Ilya Metchnikov () et à Jules Bordet () pour leurs travaux
respectifs sur la phagocytose et sur le système du complément, les onze autres
immunologistes lauréats du prix se sont vus récompenser pour leurs travaux
sur les composants spéciques du système immunitaire.
. Homann, J.A., Kafatos, F.C., Janeway, C.A. and Ezekowitz, R.A. . Phylogenetic
perspectives in innate immunity. Science , -.
. Dans un article écrit en pour un numéro de Scientic American dédié au système
immunitaire, Avrion Mitchison décrivait brièvement le système immunitaire inné
comme “primitive and dumb”, ce qui peut se traduire par primitif et bête! (Mitchison,
A. . Will we survive? Scientic American : -)
. En , au plus fort des recherches sur le système immunitaire adaptatif, une période
baptisée «nouvelle ère immunologique» par le lauréat du prix Nobel de Physiologie ou
Médecine , Sir Franck Macfarlane Burnet (Table ), celui-ci prédit que l’immuno-
logie se retrouverait rapidement à court de problèmes à résoudre. Plus tard, en ,
Niels Jerne, prix Nobel de Physiologie ou Médecine en pour ses travaux sur l’im-
munité adaptative (Table ), annonça que l’immunologie serait complètement résolue
dans les ans à venir. Notez que l’histoire voudrait que ce dernier ait refusé que des
séminaires sur les macrophages, cellules clés de l’immunité innée, soient organisés à
l’Institut d’Immunologie de Bâle! (Anderson, W., Jackson, M. and Rosenkrantz, B.G.
(). Toward an unnatural history of immunology. J. Hist. Biol. ():-).

: , …
Table 1. Prix Nobel de Physiologie ou Médecine couronnant
des recherches en immunologie.
Date Nom du(des lauréat(s) Intitulé du prix* Commentaires
Ilya I. Metchnikov et
Paul Ehrlich
En reconnaissance de leurs
travaux sur l’immunité
Découverte de la
phagocytose
Jules Bordet Pour ses découvertes relatives à
l’immunité
Découverte du
complément (alors
appelé Alexine)
Sir F. Macfarlane
Burnet et Peter B.
Medawar
Pour leur découverte de la
tolérance immunologique acquise
La non réponse
immune spécique
Baruj Benacerraf, Jean
Dausset et George Snell
Pour leurs découvertes concernant
les structures génétiquement
déterminées à la surface des
cellules qui régulent les réactions
immunologiques
Le Complexe Majeur
d’Histocompatibilité
(CMH appelé HLA
chez l’homme pour
Human Leucocyte
Antigen)
Niels Jerne,
Georges J.F. Köhler et
Cesar Milstein
Pour les théories concernant la
spécicité dans le développement
et le contrôle du système
immunitaire et la découverte du
principe de production des
anticorps monoclonaux
Susumu Tonegawa Pour sa découverte des principes
génétiques à l’origine de la
diversité des anticorps
Peter Doherty et Rolf
M. Zinkernagel
Pour leur découverte concernant
la spécicité de la défense
immunitaire cellulaire
Rôle du CMH dans la
présentation de
peptides aux lympho-
cytes T
Jules Homann et
Bruce Beutler
Ralph Steinman †
Pour leurs travaux sur le système
immunitaire inné
Pour ses travaux sur le système
immunitaire adaptatif
Découverte des
Toll-Like Receptors
(TLRs)
Découverte des cellules
dendritiques (DC)
* source: http://www.nobelprize.org/nobel_prizes/medicine/laureates
En dépit de cette dichotomie dogmatique qui séparait l’immunité innée
et adaptative, il a été démontré que ces deux parties du système immunitaire
sont intimement liées et qu’elles fonctionnent en synergie pour cibler ecace-
ment l’infection tout en épargnant les tissus de l’hôte. L’idée d’un système
immunitaire inné rudimentaire comparé à une réponse adaptative «plus com-
plexe» n’est ainsi plus en vigueur. Le système immunitaire des vertébrés est
 6
7
8
9
10
11
12
13
14
15
16
17
18
6
7
8
9
10
11
12
13
14
15
16
17
18
1
/
18
100%