27/09/13 BINET Lauren L3 Appareil respiratoire Pr

APPAREIL RESPIRATOIRE – Physiologie du contrôle ventilatoire
27/09/13
BINET Lauren L3
Appareil respiratoire
Pr. JAMMES
8 pages
PHYSIOLOGIE DU CONTROLE VENTILATOIRE
Le diapo du cours a été publié sur le groupe de la promo. Les illustrations et graphiques n'étant donnés qu'à
titre indicatif, ils ne sont pas à apprendre. Toutes les explications en rapport sont dans ce ronéo.
1/8
Plan
A. Automatisme respiratoire et « centres respiratoires »
I. Le centre respiratoire « classique »
II. Le centre respiratoire « moderne »
III. Genèse du rythme ventilatoire : 3 théories
B. Modulation de la ventilation par les informations périphériques
I. Les afférences vagales
II. Les messages d'origine circulatoire
III. Les afférences musculaires : muscles respiratoires et muscles squelettiques
C. Adaptations respiratoires à l'exercice
I. La cascade de l'oxygène
II. Indication des épreuves d'efforts cardiorespiratoire et métabolique
III. Les modifications normales lors d'un effort

APPAREIL RESPIRATOIRE – Physiologie du contrôle ventilatoire
A- Automatisme respiratoire et « centres respiratoires »
I. Le centre respiratoire « classique »
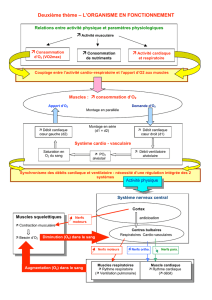
On peut volontairement être en apnée ou en hyperventilation, ceci grâce au cortex cérébral.
L'automatisme ventilatoire siège dans le tronc cérébral (bulbe rachidien ++).
Il est responsable de l'activité respiratoire.
Pour le prouver, on fait une expérience :
On réalise une section entre le mésencéphale et le tronc cérébral : la respiration est normale (eupnée). En
revanche si on sectionne en-dessous du bulbe (= moelle allongée) : la respiration s'arrête (apnée).
Le contrôle se situe donc dans le tronc cérébral, en-dessous du mésencéphale.
II. Le centre respiratoire « moderne »
Il existe une localisation précise des neurones propriobulbaires et des voies bulbospinales commandant les
motoneurones phréniques, intercostaux et abdominaux.
Les neurones propriobulbaires sont entièrement inclus dans le tronc cérébral, à la différence des neurones
bulbospinaux.
On retrouve 2 populations de neurones propriobulbaires :
–Le groupe respiratoire dorsal (DRG) qui commande le nerf phrénique (le diaphragme représente 90%
des mouvements respiratoires) → important +++
–Le groupe respiratoire ventral (VRG) qui commande les muscles accessoires (muscles intercostaux et
abdominaux) qui suppléent le diaphragme (chant, cri...) en dehors de la respiration calme.
Les neurones ont une activité automatique.
Les neurones respiratoires propriobulbaires commandent, via les voies bulbospinales, les motoneurones spinaux
(de la moelle) :
–Nerfs phréniques (C3 à C5): nerfs fragiles essentiels à la vie : ils descendent jusqu'au diaphragme
(≈ 60 cm de long pour 1,5 à 2 mm de diamètre d'où sa fragilité) le long du médiastin et du cœur et se
projettent en parapluie sur chacune des coupoles.
Le nerf phrénique est soumis à pathologies, comme les cancers du médiastin, un lymphome par
exemple peut le comprimer.
–Nerfs intercostaux (T1 à T12)
–Nerfs abdominaux (L1 à L4) tous expiratoires.
A leur tour, les nerfs commandent les muscles respiratoires via les voies motrices.
III. Genèse du rythme ventilatoire : 3 théories
Première théorie : il y a une dépolarisation spontanée du potentiel de repos permise par la présence de
neurones « pace-makers »
Deuxième théorie : il y a un réseau neuronal (neurones inspiratoires (I), expiratoires (E) et interphasiques (E-I
ou I-E)), un rythme s'installe et repose sur les oscillations spontanées de ce réseau.
Troisième théorie : le mécanisme du off-switch = interruption de l'inspiration par des messagers d'origine
pulmonaire. Lorsqu'on respire, des informations viennent du poumon, transitent par le nerf vague (X) et
inhibent l'activité respiratoire : plus le poumon se gonfle, plus il se distend et plus les informations sont
importantes. L'inspiration devient de plus en plus courte donc la fréquence respiratoire augmente.
Chez l'individu sous anesthésie, il n'y a pas d'activité comportementale et le mécanisme off-switch est très
important.
Selon le prof, ces 3 phénomènes coexistent.
2/8

APPAREIL RESPIRATOIRE – Physiologie du contrôle ventilatoire
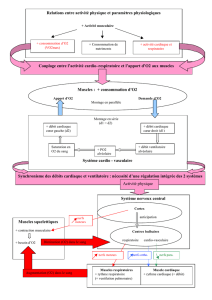
B. Modulation de la ventilation par les informations périphériques
L'activité des neurones respiratoires propriobulbaires est modulée par des informations périphériques. Ces
dernières viennent du poumon et du cœur (via le nerf vague sensitif ++), du larynx (phonation, via le nerf
laryngé supérieur).
Le NTS (Nucleus Tractus Solitarius ou faisceau solitaire nucléaire) conduit les informations végétatives : il
reçoit 90% des informations périphériques provenant de chimiorécepteurs artériels, qui modulent l'activité
des neurones respiratoires automatiques en projetant sur DRG et VRG.
Les muscles respiratoires et squelettiques ne passent pas par le NTS, ils projettent directement sur l'appareil
respiratoire dorsal ou passent par le cervelet. Ils renseignent sur l'efficacité des muscles respiratoires et sur
l'activité physique.
On démontre le rôle fondamental des afférences respiratoires (= informations périphériques) dans l'ajustement
de la commande ventilatoire par une expérience sur un chien.
- Lorsque le chien est éveillé, la respiration est normale. On enregistre la fréquence respiratoire à 14-15/min.
- Le chien s'endort, il y a une chute de la fréquence respiratoire à 10/min. → Les centres du comportement
jouent donc un rôle important.
- Lors d'une opération, les nerfs vagues du chien sont placés sous la peau. On bloque ces nerfs en appliquant
des glaçons sur la peau, la respiration ralentit alors considérablement (4-5/min).
- Le chien respire de l'O2 pur : cela bloque la décharge des récepteurs artériels, la fréquence respiratoire diminue
encore.
- Le chien est perfusé avec des bicarbonates : le pH augmente, la fréquence respiratoire diminue → les
chimiorécepteurs centraux sont sensibles au pH.
L'animal est toujours vivant mais respire à 2 mouvements par minute : il est hypoxique et hypercapnique.
==> Ceci montre qu'il existe bien un automatisme respiratoire car si on supprime les informations
périphériques, on ne supprime pas totalement la respiration, il persiste un rythme ventilatoire primaire.
Cependant, à la différence de l'automatisme cardiaque, il ne permet pas de maintenir longtemps l'individu en vie
(ce qui est une cause de la mort subite du nourrisson par exemple) et doit donc être stimulé par les informations
périphériques.
I. Les afférences vagales
Le nerf vague (6 mm de diamètre) alimente tous les organes du thorax et 70% des organes du tube digestif. Il
est à la fois moteur et sensitif et comporte une voie montante et une voie descendante.
Le nerf vague a une particularité : il n'y a pas de ganglion moteur mais un volumineux ganglion sensitif sous la
mâchoire: le ganglion plexiforme (ou nodal) qui contient tous les noyaux des neurones sensitifs.
Le nerf vague descend le long de la trachée, du péricarde, se divise au hile du poumon puis continue le long du
tube digestif jusqu'au côlon transverse.
Les afférences vagales respiratoires proviennent des mécanorécepteurs du vague, des récepteurs d'irritation et
des fibres C.
Les mécanorécepteurs du vague présents dans les grosses voies aériennes (= trachée et bronches, il n'y PAS de
mécano-sensibilité dans les alvéoles) transmettent des informations. Ces récepteurs pulmonaires sont sensibles
à l'étirement (PSR = Pulmonary Stretch Receptor) et projettent sur le faisceau solitaire (NTS).
Ils déchargent à chaque inspiration et arrêtent de décharger à l'expiration. Plus les poumons se gonflent, plus la
décharge est importante.
La décharge est synchrone et correspond à l'étirement des parois trachéobronchiques. Ces récepteurs
renseignent sur l'amplitude du volume courant et sur la fréquence respiratoire.
3/8

APPAREIL RESPIRATOIRE – Physiologie du contrôle ventilatoire
En réponse à ces informations, il y a une interruption de la décharge des neurones inspiratoires (off-switch) →
C'est le réflexe de Breuer et Hering. Il y a une inhibition inspiratoire, plus le volume pulmonaire est
important, plus l'inspiration est inhibée. Ceci assure le contrôle de la fréquence respiratoire.
Le deuxième type de récepteurs transmet des informations sur la composition chimique et la température des
gaz ventilés, ce sont des récepteurs dits d'irritation (détectent air FROID)
Ils sont sensibles à l'irritation du larynx et des bronches et déclenchent la toux et l'éternuement. Ils ne jouent pas
de rôle dans la physiologie.
Les fibres amyéliniques (fibres C) sont situées partout, y compris dans les alvéoles. On trouve des fibres C
trachéobronchiques et des fibres C alvéolaires. Elles renseignent aussi sur la composition et la température des
gaz inhalés.
90% des informations qui viennent des poumons transitent par ces fibres, elles représentent donc 90% des
voies sensitives du nerf vague pulmonaire.
Les fibres C mesurent beaucoup de paramètres : l'inflammation des parois trachéobronchiques, la distension
des alvéoles, la pression dans la circulation pulmonaire (détecte l'hypertension artérielle pulmonaire ou HTAP),
le refroidissement des voies aériennes, les toxiques, le flux de CO2 qui sort des poumons (activité musculaire
à l'exercice).
Le niveau de vigilance est très important dans le contrôle ventilatoire par le nerf vague car les informations
sont plus nombreuses lorsque l'on est endormi. Chez l'Homme endormi (ou anesthésié), l'inhibition respiratoire
est plus grande. En effet, lorsque l'on est éveillé, le réflexe off-switch est beaucoup moins important. Plus le
volume pulmonaire est important, plus le réflexe est diminué.
II. Les messages d'origine circulatoire
Ils renseignent sur la composition en O2 (PaO2) et en CO2 (PaCO2) du sang artériel.
Les chimiorécepteurs artériels sont présents dans les bifurcations carotidiennes (ce qui pose un problème en
vieillissant car l'athérosclérose se localise d'abord principalement au niveau du sinus carotidien → le signal
chimiosensible diminue), sur la crosse aortique et l'aorte thoracique.
Au niveau du sinus carotidien, ce sont des amas cellulaires, des glomi qui sont extériorisés de la paroi (à la
différence des barorécepteurs) et ont une vascularisation propre. C'est le nerf glosso-pharyngien (IX) qui
transmet l'information (rôle +++).
Les informations provenant de la crosse aortique empruntent la voie du nerf vague.
Ces messages sont envoyés au niveau du tronc cérébral et vont contrôler à la fois les voies respiratoires, les
nerfs phréniques et les neurones commandant l'activité sympathique.
2 types de neurones sont concernés : ceux du groupe respiratoire dorsal et ceux du groupe respiratoire ventral.
Quand la PaO2 baisse, il y a donc une hyperventilation.
Au niveau du tronc cérébral, des neurones contrôlent l'activité sympathique et donc l'activité cardiaque.
L'activation des chimiorécepteurs artériels augmente la décharge des neurones inspiratoires et expiratoires
(chimioréflexe respiratoire) et stimule la commande sympathique cardiaque et vasculaire (chimioréflexe
cardiovasculaire) → augmentation du débit cardiaque.
➢Ces chimiorécepteurs artériels sont imparfaits car il faut attendre que la PaO2 diminue fortement pour
qu'ils déchargent. En effet, la réponse ventilatoire à l'hypoxie est non linéaire car les récepteurs ont
un seuil de décharge : 75 mmHg, l'hypoxie est alors déjà bien entamée. Dans l'idéal, il faudrait qu'ils
déchargent dès que la PaO2 descend en-dessous de 90 mmHg.
L'organisme se défend donc mal contre l'hypoxie.
4/8

APPAREIL RESPIRATOIRE – Physiologie du contrôle ventilatoire
➢A l'inverse, ces chimiorécepteurs sont très sensibles à l'hypercapnie. Ils n'ont pas de seuil et détectent
une variation de 2 mmHg. La décharge est proportionnelle à la PaCO2. La relation entre degré
d'hypercapnie et réponse ventilatoire est linéaire.
➔Les mêmes récepteurs détectent l'hypoxémie et l'hypercapnie.
Chimiosensibilité centrale
Des neurones du tronc cérébral situés au niveau du plancher du IVème ventricule détectent l'hypercapnie et
l'acidose grâce à leur sensibilité à des agents chimiques passant dans le sang circulant et le liquide céphalo-
rachidien mais ne détectent PAS l'hypoxie (l'hypoxie ne stimule QUE les récepteurs périphériques).
Lors d'une hypercapnie et/ou d'une acidose, ils entraînent une hyperventilation et une augmentation du débit
cardiaque.
L'hypoxie périphérique stimule la ventilation, l'hypoxie centrale régule l'activité respiratoire (arrêt respiratoire).
Chimiosensibilité artérielle et centrale chez l'Homme
On mesure le volume expiratoire en fonction du temps en faisant respirer soit du CO2, soit de l'O2 pur au sujet.
–Après une bouffée de CO2, le volume expiratoire augmente en quelque secondes : il y a une réponse
globale, la chimiosensibilité est artérielle (périphérique) PUIS centrale.
–Une bouffée d'O2 pur bloque les chimiorécepteurs artériels, puis au bout de 10 à 15 secondes, le sujet
commence à hyperventiler : mise en jeu des chimiorécepteurs centraux.
Les barorécepteurs artériels assurent également un contrôle essentiel lors de la respiration.
➔Il y a donc 2 systèmes de détection des anomalies des gaz du sang.
III. Les afférences musculaires : muscles respiratoires et muscles squelettiques
Sensibilité musculaire et tendineuse
Il existe des fibres qui ne détectent que les modifications mécaniques : ce sont les organes tendineux de Golgi.
Ils informent sur le mouvement (marche, étirement...) et sont situés au niveau de la jonction fibres musculaires/
tendons.
Il existe également des fuseaux neuromusculaires, constitués de fibres musculaires et de terminaisons
nerveuses libres.
Ces 2 types de structures fournissent des renseignements en permanence.
Les terminaisons nerveuses libres de type III et IV représentent plus de 40% des informations qui mesurent le
mouvement et l'activité biochimique des muscles : ce sont des métaborécepteurs.
Les fuseaux neuromusculaires contiennent des fibres innervées par des motoneurones dynamiques ou statiques.
La partie centrale du fuseau est non contractile et encapsulée, c'est la partie sensible. Ils contiennent des fibres
musculaires et des fibres nerveuses → organes sensitifs et contractiles.
Leur décharge est réglée par le système nerveux central. La décharge mesure le mouvement mais est amplifiée
ou diminuée par la commande centrale.
Il existe des fuseaux neuromusculaires dynamiques et statiques : ils détectent avec un seuil très bas la moindre
contraction musculaire :
–Fuseau dynamique décharge lorsque vitesse de contraction est très importante,
–Fuseau statique est sensible à l'étirement prolongé
–Fuseau tendineux de Golgi sensible au moindre étirement du tendon.
5/8
 6
7
8
6
7
8
1
/
8
100%