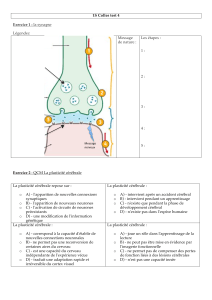
Plasticité nerveuse - Plan et documents de cours

E54PS3 : Psychophysiologie des comportements : Plasticité nerveuse et mémoire
Page 1
Documents de cours destinés aux étudiants de la licence de
psychologie L3S5 – 2015-2016
Plasticité nerveuse et mémoire
imechaly@univ-montp2.fr
Références bibliographiques

E54PS3 : Psychophysiologie des comportements : Plasticité nerveuse et mémoire
Page 2
PLAN DU COURS
Plasticité nerveuse et mémoire
I. Introduction
1. Le tissu nerveux : un réseau extraordinairement complexe
2. La plasticité cérébrale : Définition
3. Neuroplasticité et mémoire
4. Contributions majeures dans le domaine de la plasticité
II. Plasticité et Développement
A. La construction des circuits nerveux en sept étapes
1. Neurogénèse
2. Migration cellulaire
3. Différentiation cellulaire
4. Maturation
a. croissance dendritique et axonale
b. Synaptogénèse
c. Mort cellulaire
d. Myélogénèse
B. Modifications des circuits cérébraux sous l’effet de l’expérience
1. La notion de période critique
2. Privation visuelle et développement visuo-moteur
3. Le Postulat de Hebb
4. Rôle du calcium dans la plasticité dépendante de l’activité
5. Développement et environnement enrichi
III. Plasticité post-développementale
A. Le rôle des synapses dans l’apprentissage et la mémoire
1. Plasticité synaptique à court terme et mémoire non déclarative
a. L’habituation
b. La sensibilisation
c. Rappel sur le fonctionnement de la synapse
d. Bases moléculaires de la plasticité à court terme chez l’aplysie
e. La potentialisation post-tétanique chez le mammifère
2. Plasticité à long terme et mémoire déclarative
a. Les synapses de L’hippocampe
b. Les bases moléculaires de la PLT
c. PLT et mémoire déclarative
d. La dépression à long terme
e. Changements moléculaires et structuraux dans la PLT tardive

E54PS3 : Psychophysiologie des comportements : Plasticité nerveuse et mémoire
Page 3
B. Plasticité dans le cortex cérébral adulte
IV. Les circuits de la mémoire
1. Théorie des assemblées cellulaires de Hebb
2. Le cas du patient H.M
3. Structures cérébrales impliquées dans la mémoire déclarative
4. Structures cérébrales impliquées dans la mémoire non déclarative
Quelques diapos de cours et documents
I. Introduction

E54PS3 : Psychophysiologie des comportements : Plasticité nerveuse et mémoire
Page 4
II. Plasticité et développement
A. La construction des circuits nerveux
Les principales étapes de l’ontogénèse cérébrale
Le cortex commence à se former vers l’âge de 6 semaines (post-fécondation), la
neurogénèse étant achevée en grande partie vers l’âge de 20 semaines. La migration
neurale et la différenciation cellulaire démarrent vers l’âge de 8 semaines. Elles sont
en grande partie terminées à l’âge de 29 semaines. La maturation des neurones qui
inclut la formation des dendrites et des axones, commencent vers l’age de 20
semaines et continue bien après la naissance. La masse du cerveau comme celle du
corps croissent rapidement et en parallele, pendant la periode prénatale. (Cerveau et
comportement, Kolb et Wishaw)

E54PS3 : Psychophysiologie des comportements : Plasticité nerveuse et mémoire
Page 5
La migration cellulaire
Production programmée de différents types de neurones à différents moments à partir des
cellules précurseurs ou progénitrices en cours de division du cortex cérébral du cerveau d’un
mamifère. Près d’une face du neuroépithelium cortical, les cellules progénitrices se divisent
de façon répétitive comme une cellule souche, pour produire des neurones. Les neurones
migrent vers l’exterieur en direction de la face opposée de l’épithélium le long de la surface
des cellules radiales de la glie. Les neurones nés les premiers s’établissent le plus près de
leur lieu de naissance tandis que les neurones nés plus tard migrent et les dépassent pour
s’établir plus loin. Des générations succéssives de neurones occupent ainsi différentes
couches du cortex et ont différents caractères intrinsèques selon leur date de naissance.
(Biologie Moléculaire de la cellule, Alberts et al.)
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
1
/
35
100%