Neuroendocrinologie (en )

NEUROENDOCRINOLOGIE
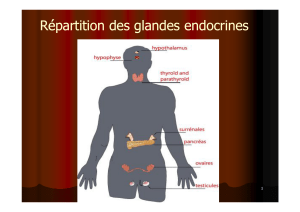
L'AXE HYPOTHALAMIQUE
Sécrétion d'hormone par un neurone, qui sont des cellules excitables et
qui libèrent des neurohormones.
L'axe hypothalamus hypophysaire se trouve dans le cortex cérébral,
l'hypophyse étant en dessous de l'hypothalamus. Ils sont reliés tous les deux au
reste du cortex par la tige hypophysaire (ou pituitaire)
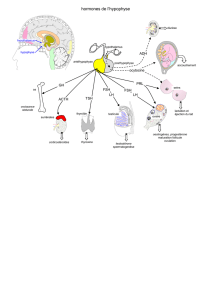
L'hypophyse est constituée de 3 parties :
Le lobe nerveux (pars nervosa) qui correspond à l'hypophyse ou lobe
postérieur. Cette partie libère la vasopressine (ADH) et l'ocytocine. ADH cible
les cellules du tube collecteur au niveau du rein et les artères (pour la
pression). L'ocytocine cible les muscles utérins (contraction), les canaux
lactatifs (éjection du lait). Elle permet également l'éjaculation et certainement
d'autres mécanismes mais son rôle est mal connu chez l'homme.
Le lobe antérieur est composé de 2 parties : la pars intermedia et la pars
distalis. Ce lobe libère ACTH, LH/FSH, TSH, GH et Prolactine
*) La prolactine agit sur les glandes mammaires (synthèse de lait) et sur le
système immunitaire.
*) La GH (Grow Hormon) agit sur le foie (favorisation du métabolisme) et sur les
os
*) La TSH (Thyroïd Stimulating Hormon) agit sur la thyroïde qui en retour agit sur
le métabolisme en général
*) La LH/FSH (Hormones gonadotropiques) agit sur les gonades :
Chez la femme : FSH stimule la phase folliculaire et les cellules de la
granulosa. LH sur les cellules de la thèque et du corps jaune.
Chez l'homme : FSH agit sur les cellules de Sertoli. LH sur les cellules de
Leydig pour la synthèse de testostérone (quia git sur le métabolisme, voix, poils,
muscles,...)
*) L'ACTH (adénocorticotropine homorn) agit dans la production de cortisol au
niveau des glandes corticosurrénales. Lors d'un stress le cortisol agit sur la
néoglucogenèse. Il agit aussi sur la diminution du système immunitaire lors
d'inflammation. Le cortisol agit au niveau fœtal au moment de la parturition
(engagement de celle ci)
On trouve 5 types de cellules ce qui explique pourquoi LH et FSH sont
ensemble. Les cellules gonadotropes (LH/FSH), somatotrope (GH), Cellules à
prolactine, cellules corticotropes (ACTH) et cellules thyroïcotropes (TSH).
Le lobe intermédiaire synthétise une énorme préhormone la POMC qui
donne P ( pou pré hormone), O pour les endorphines, M pour MSH et C pour ACTH
(mais on arrive pas vraiment à la quantifier)
MSH (mélanocyte stimulating hormon) permet un changement de la
pigmentation (ce qui n'est pas le cas chez nous, où seul le soleil colore notre
peau, on ne peut pas foncer ou éclaircir volontairement notre peau).
Chez les Amphibiens elle permet de faire varier la couleur de la peau selon
le milieu où ils se trouvent (plus ou moins foncée par la migration au sein des

mélanocytes soit vers la membrane (foncement) soit vers le centre de la cellule
(éclaircissement))
2 origines pour la mise en place des cellules de l'hypophyse.
RATHKE 1835 (poly page 1)
La formation d'un diverticule qui grandit puis se détache du stomodéum pour
venir se fixer sous le plancher (infundibulum) de l'hypothalamus qui lui aussi émet
un diverticule, permet de « construire » l'axe.
Chez le rat, la poche de RATHKE conserve une cavité appelée fente
hypohysaire qui permet de délimiter les 2 parties du lobe. Chez l'homme cette
distinction est difficile sauf par marquage car la cavité n'est pas présente.
Le lobe nerveux contient les terminaisons de neurones neurosécréteurs
(il n'y pas de cellules glandulaires) responsables de la sécrétion d'ADH et
Ocytocine.
Cependant les expériences de KAWAMURA (en 1998) les 2 parties de
l'hypophyse ont une origine nerveuse. Il a greffé au niveau de territoires
présomptifs des cellules pigmentées sur un œuf fécondé non pigmenté.
Les bords du tube neural (crêtes neurales) constituent les lobes
antérieurs de l'hypophyse et ne viennent pas du stomodénum.
Pit : cellules olfactives
Br : ventricule
Ht: donnera le futur hypothalamus
Tube neural pour le lobe nerveux
Crête neural pour le lobe antérieur
Relation entre le diverticule et le stomodénum par le plancher et le
diverticule. Si l'on empêche cette intéraction physique, on observe pas de
formation de l'hypophyse, pareil si cela est fait entre le toit et le stomodénum.
1er facteur de croissance BMP4
provenant du plancher de
l'hypothalamus et doit pouvoir
migrer vers la poche de RATHKE.
Ce facteur permet la synthèse
d'un facteur Trf1
2cd facteur de croissance FGF8
provient du toit du stomodénum
et migre dans la poche et permet
la production d'autres facteurs
permettant la fin de la formation
de l'hypophyse.
(poly page 3) Tableau n°1
Large présence de somatotrope et gonadotrope 60 à 75% du nombres totale de
cellules de l'adénohypophyse.
C'est la taille des grains de sécrétions qui doit permettre de distinguer les
types cellulaires. C'est plutôt grâce à des expériences telle la castration qui
provoque une hypertrophie des cellules gonadotropes. Sur un rat castré à qui on

injecte des broyas de testicules on remarque que les cellules reviennent à une
taille normale. Mise en évidence d'un feed back négatif (rétrocontrôle) des
hormones des gonades sur les cellules gonadotropes.
Pour les autres cellules c'est pus difficile du fait de leur action globale,
cependant une carte des types des cellules a pu être faite.
Cette hypophyse régule l'organisme ou d'autres glandes endocrinienne
mais elle doit également être régulée. C'est le rôle de l'hypothalamus.
Fonction du lobe nerveux dépend bien de l'hypothalamus car on trouve des
molécules qui activent :
libérines (axe mélanotrope (lobe intermédiaire) il possède une innervation
dopaminergique)
ou qui inhibent :
Statines DA, GABA issues de l'hypothalamus et 5-HT issue du noyau raphé
Axe gonadotrope : LH-RH (libérine) ou Gn-RH
Augmentation et libération de LH (principalement). On ne connait pas de
stratines à ce jour.
Axe corticotrope : CRH (libérine) agit sur ACTh
Axe somatotrope : GRH (libérine) qui agit sur GH et SRIF (statine)
Axe thyroïdotrope : TRH (libérine) TSH
Axe lactotrope : on ne connait rien de bien spécifique
TRH, neurotensine on aune légère stimulation sur la prolactine.
Inhibition de ces cellules par le PIF (Prolactine Inhibition Factor) en fait la
dopamine par l'hypothalamus.
Les axes somatotrope et lactotrope sont différents des autres par leur cible
(pas de cellules endocriniennes visées). Les autres sont inhibés par le feed back
négatif des hormones produites.
(poly page 4 figure 1) On s'intéresse à la formation de l'hypothalamus, donc du
devenir du diencéphale. Courbures du cerveau pour occuper la cavité crânienne et
déjà au bout de 2 mois le cerveau est quasiment définitif. Il s'hypertrophie et se
retrouve à l'intérieure de l'encéphale.
L'hypothalamus se trouve entre le chiasma optique et la partie
postérieure du noyau arqué.
Les ventricules III et IV sont assez petit. L'hypothalamus se trouve en plein sur le
plan de symétrie du diencéphale.
(poly page4 figure 5)
Noyau paraventriculaire collé au ventricule III.
Noyau médian est composé de 2 morceaux, le médian et le ventromédian.
Au dessus on trouve l'aire hypothalamique latérale. (dans une aire les
neurones sont moins ramassés sur eux mêmes).

Noyau supraoptique (en fonction de sa position très proche au chiasma
optique).
Noyau suprachiasmatique collé au chiasma optique
Noyau pré optique
Corps mammillaire
Tous ces noyaux sont pairs, c'est à dire qu'ils sont au nombre de deux, en
symétrie par rapport au plan du diencéphale.
Le seul noyau qui est impair est le noyau arqué (plancher du ventricule
III)
L'hypothalamus est une petite structure limité par le noyau préoptique et le corps
mammillaire dorseo-ventralement et le noyau arqué et l'aire hypothalamique.
(poly page 6)
Si on prend la région pré optique. Sa première fonction est de réguler la
reproduction : noyaux secrétant GnRH, eux mêmes recevant des informations
afférentes les régulant. Elle joue une fonction de thermorégulation par la
régulation des phénomènes de thermolyse. Si l'on détruit ces 2 noyaux par
électrocoagulation on observe une incapacité à percevoir l'augmentation de la
température corporelle.
Noyaux suprachiasmatiques : Ils régulent les rythmes saisonniers et
circadiens en étant directement liés à la rétine. Ce noyau est du coup renseigné
par l'œil sur la période de la nuit. Il analyse via l'épiphyse également la durée des
phases diurne et nocturne (la reproduction étant liée à la photopériode et à
l'épiphyse). Soit reproduction au début du printemps (majeur partie des animaux)
soit à l'automne (pour les Cervidés)
Noyaux périventriculaires: Fonction endocrine CRH, TRH et somatostatine
Noyaux supraoptiques : noyaux à ocytocine et vasopressine (équilibre
hydrique et reproduction). Région sensible à la pression osmotique grâce à des
osmorécepteurs.
Noyaux paraventriculaires : Moins
sensible à la pression osmotique mais on
trouve des neurones à vasopressine et
ocytocine. Régulation du stress (neurones à
CRH et TRH).
Ces noyaux sont divisés en 2 parties :
La CRH est libérée dans l'éminence médiane
Certains neurones de la zone parvicellulaire
synthétisent aussi de l'AVP et ocytocine
considéré comme des peptides accessoires.
D'autres peuvent synthétiser également de
la neurotensine.

Noyaux ventromédian :
neurones à CRH et TRH. Rôle
important dans la prise
alimentaire. Sans ces noyaux
on remarque une hyperphagie
(perte de la sensation de
satiété)
Aire hypothalamique
latéral : Perte de la sensation
de faim. Hypophagie. Le
problème d'être dans une aire
résulte de la difficulté à savoir
quels sont les neurones responsables. Cette zone synthétise le neuropeptide Y
également synthétisé par le noyau arquée.
Noyau arquée : neurones à neuropeptides Y (NPY), POMC (pro-
opiomélanocortine précurseur à l'ACTH), CART (cocaïne and amphetamine
regulated transcript) et l'AgRP (agouti-related protein). POMC et CART entraînent
une inhibition de la faim. Ils sont anorexigènes.
La barrière hématoencéphalique n'est pas présente dans la partie basale du
noyau arquée. NPY et AgRP sont inhibés lors de la prise alimentaire par des
facteurs périphériques et également par le psychisme (dans le cas d'anorexie ou
par la volonté).
Les hormones hypothalamiques qui régulent la croissance se situent dans
une zone proche de celle de la prise alimentaire.
L'hypothalamus postérieur : joue dans la thermorégulation par les
processus de thermogenèse. Ces variations captées par la moelle épinière. Régule
aussi plus ou moins l'alternance veille/sommeil, agressivité, fuite.
Hypophyse antérieur : Chez les Poissons les axones libèrent directement
au niveau du lobe antérieur les hormones dans la circulation sanguine. A partir
des Lissamphibiens les neurones hypothalamiques utilisent un relais (le système
porte) et projettent leurs neuromédiateurs dans l'éminence antérieure.
Chez les Poissons, l'origine des neurones régulant l'activité de l'hypophyse a
été déterminé grâce à des colorations. On dépose un bout de colorant et on
attend quelques jours et l'on fait des coupes du cerveau. Ce sont les neurones de
l'hypothalamus par innervation directe qui régulent l'activité du lobe nerveux (cas
de tous les Vertébrés) et sur l'hypophyse (cas des Poissons) au niveau des cellules
gonadotropes.
Chez les Vertébrés, ces neurones n'envoient pas leurs statines et libérines
directement. Ils les libèrent dans une zone très vascularisée (système porte) où
d'autres neurones récupèrent l'information et agissent sur les cellules
gonadotropes de l'hypophyse antérieure.
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%
![Découverte d un nouveau centre cérébr[...]](http://s1.studylibfr.com/store/data/001261824_1-044b689d1e2faad91148811640c2eb34-300x300.png)



![Découverte de 30 petits neurones qui en[...]](http://s1.studylibfr.com/store/data/001768997_1-600bf3836f939b795834ee592d4e2e5f-300x300.png)