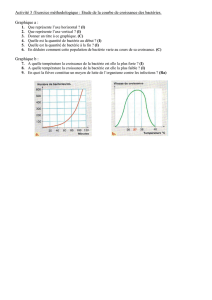
III - Interactions entre l`Homme et les

Les bactéries
I) Introduction
Depuis 1941 on utilise un antibiotique, la pénicilline, mais une mauvaise utilisation
(excessive) est à l’origine de résistances (que l’on redoute depuis 1945). En effet on
sélectionne les bactéries résistantes et les antibiotiques n’ont plus aucune efficacité.
D’autres antibiotiques ont été développés :
Pénicilline paroi (Gram + ou Gram -).
Sulfamide synthèse de l’acide folique.
Chloramphénicol synthèse protéique.
Depuis une vingtaine d’années, on ne trouve plus de nouvelles cibles pour les antibiotiques
(ADN girase, élongation des ribosomes…). On peut essayer un criblage de molécules au
hasard pour découvrir lesquelles auraient une action antibiotique, mais ce n’est pas très
efficace.
Pour combattre les pathogènes, il faut savoir comment l’hôte réagit aux pathogènes. C’est le
meilleur moyen pour trouver de nouvelles clés de combat contre les microbes (antibiotiques,
substances permettant de conférer une résistance proche de la vaccination…). Une nouvelle
discipline a donc émergé : la microbiologie cellulaire.
Les agents pathogènes sont des microbes présents partout, mais ayant acquis des mécanismes
pour interagir avec l’hôte. L’homme possède 1013 cellules, mais il a aussi 10 fois plus de
bactéries et 100 fois plus de virus. Ceci s’explique par la rapidité de la reproduction
bactérienne (20 minutes – 1 heure) par rapport à la division des cellules humaines.
La plupart des bactéries et virus existent donc en « bonne entente » avec nous, mais de temps
en temps des mutants développent des mécanismes en leur faveur, agressant l’hôte.
Nous allons voir 3 exemples d’espèces de bactéries :
- E. coli : très connu
- Shigella flexneri : même génome que E. coli, mais avec un plasmide de virulence en plus. Il
provoque des shigelloses (diarrhées profuses pratiquement continues), et est fréquent dans les
zones intertropicales (du Pakistan au Bengladesh). Les enfants atteints se déshydratent et
meurent rapidement. Cette maladie est traitée avec des antibiotiques.
- Salmonella enteritica : même génome que E. coli, mais avec des îlots de pathogénicité
(morceaux d’ADN insérés dans le génome). C’est plus difficilement curable.
Ces 3 espèces sont très proches, mais quelques petites différences confèrent de nouveaux
caractères à la bactérie.
II) Mode d’action
Un agent pathogène agit de la manière suivante :
- coloniser l’hôte (adhésion à la surface des cellules, dans le cas contraire elle est éliminée par
lavage).
- trouver une niche nutritionnelle : être protégé et trouver ce qui est nécessaire à la survie
même s’il y a besoin de dégrader la source d’énergie.
- contourner les défenses de l’organisme, les éviter ou subvertir les réponses de l’hôte.
- se répliquer au détriment de l’hôte.
- sortir et se transmettre à un nouvel hôte.

Les signes et les symptômes peuvent être provoqués par le pathogène, mais sont souvent des
réactions de l’hôte :
- bactérie entéropathogène : diarrhées et dissémination
- lésions de l’herpès : dissémination
- réactions de l’hôte : fièvre, inflammation, rougeurs
Un exemple de colonisation de l’hôte est Helicobacter pylori qui colonise les villosités de
l’estomac. Elle sécrète une enzyme, l’uréase, qui produit de l’ammoniaque neutralisant
l’acidité de l’estomac. Cette bactérie est responsable d’ulcères gastriques pouvant évoluer en
cancers, et on traite par trithérapie (3 antibiotiques). 70% des personnes sont infectées dans le
pourtour méditerranéen.
Autre exemple : Staphylococcus aureus (des coques collées par 2, formant des diades). Elle
sécrète des toxines qui perforent de façon temporaire les cellules endothéliales (paroi du
vaisseau sanguin) : les bactéries accèdent alors à la matrice extracellulaire et se fixent sur le
collagène.
III) Adhésion à l’hôte et entrée dans l’hôte
Adhérer à l’hôte est une stratégie connue à toutes les entérobactéries, par exemple E. coli des
souches provoquant des troubles urinaires. Celles-ci sécrètent à leur surface des adhésines Afa
(afimbriées, poils plus petits que les fimbriés qui eux-même sont plus petits que les pilis) qui
reconnaissent le DAF (Decay accelerating factor, régulant la coagulation) cellulaire.
Les adhésines Afa, invisibles au microscope, sont mises en évidence par immunocytochimie à
l’or colloïdal.
Il existe deux mécanismes pour l’entrée des bactéries : le zipper (fermeture éclair) et le trigger
(entrée déclenchée). Celles-ci permettent d’entrer sans causer de dégâts (comme
Staphylococcus aureus qui détruit les cellules) et donc d’avoir une véritable niche
nutritionnelle : l’intérieur d’une cellule est un excellent milieu de culture !
1) Zipper (fermeture éclair)
La bactérie sécrète des molécules d’adhésion (adhésines) à sa surface, interagissant avec les
intégrines ou les cadhérines de la cellule hôte. La bactérie va, par le mouvement des
adhésines, tirer la membrane plasmique à elle, s’enroulant et pénétrant ainsi dans la cellule.
Exemple : Listeria monocytogenes qui se réplique à basse température et pousse encore mieux
dans les réfrigérateurs, causant la listériose. Voici son cycle :

2) Trigger (déclenchée)
De véritables seringues d’injection interviennent. Ce sont des appareils de sécrétion de type
III (il en existe 4 types différents), qui injectent dans la cellule hôte des molécules provoquant
la polymérisation des microfilaments d’actine, par le jeu des GTPases de la famille Rho. La
polymérisation de ces filaments va permettre à la cellule hôte d’ingérer la bactérie, celle-ci va
donc rentrer sans rien faire.
Les seringues proviennent d’une classe présente dans le génome, qui a été légèrement
modifiée : les moteurs flagellaires (cf cours du premier semestre). Le flagelle est enlevé, et le
crochet est modifié en système d’injection.
On peut citer comme exemple Shighella, Salmonella…
IV) Passage d’une cellule à une autre par action sur l’actine
Des protéines vont détourner l’activité de la cellule hôte et entraîner une polymérisation de
l’actine, ce qui permet un déplacement dans le cytoplasme de la bactérie. La membrane
plasmique peut ainsi être déformée, et la bactérie peut atteindre une autre cellule où elle
entrera par phagocytose. Les hémolysines sont capables de dégrader deux membranes sans
problème :

Ceci permet à la bactérie de passer d’une cellule à l’autre sans entrer en contact avec le
système immunitaire (pas de sortie des cellules).
La formation de ces microfilaments d’actine est due au complexe protéique Arp 2/3 et WASP
(provenant de l’hôte et non de la bactérie). La bactérie va sécréter Act A, sur laquelle se fixe
Arp 2/3 : ceci déclenche la formation de nombreux filaments d’actine. C’est un système en
cascade qui va permettre la formation de ces « comètes », les bactéries poussées par l’actine
qui se polymérise vont rejoindre d’autres cellules.
1
/
4
100%