Stabilité, variabilité des génomes et évolution

I- La reproduction et le brassage génétique
A- La reproduction sexuée et la conservation du caryotype de l’espèce
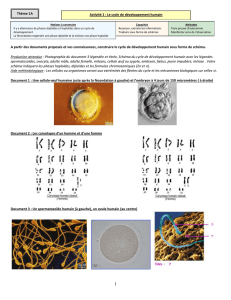
1- Une alternance de générations : les cycles biologiques
a- Chez les organismes diploïdes
Chez les mammifères comme l’Homme, les
cellules somatiques sont diploïdes (2n
chromosomes) à la différence des cellules
germinales, les gamètes, qui sont haploïdes (n
chromosomes).
Les gonades fabriquent des cellules haploïdes
grâce au mécanisme de la méiose.
La rencontre de deux cellules reproductrices
permet de former un nouvel individu. La méiose
précède de très peu la fécondation. La phase
haploïde, réduite dans le temps et l’espace, est
dominée par la phase diploïde.
b- Chez les organismes haploïdes
Chez les Champignons comme Sordaria, chaque
mycélium est une suite de cellules haploïdes. La
reproduction sexuée fait intervenir non pas des
gamètes mais deux filaments différents. Deux
cellules s’unissent pour donner une cellule unique
à deux noyaux haploïdes qui se multiplie pour
donner un organe en forme de coupe, le périthèce.
Certaines cellules de celui-ci ont leurs noyaux qui
fusionnent pour donner des cellules-œufs
diploïdes. Chaque cellule-œuf entre alors en
méiose suivie d’une mitose. On obtient 8 cellules
haploïdes ou ascospores enfermés dans un asque.
Après sa libération, une ascospore donne par
mitoses un nouveau mycélium à cellules
haploïdes.
La méiose suit donc immédiatement la
fécondation. La phase diploïde est alors réduite à
la cellule-œuf (zygote).
Stabilité, variabilité des
génomes et évolution

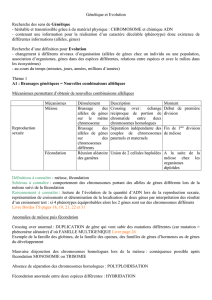
2- La méiose et la fécondation sont complémentaires
a- Le mécanisme de la méiose
La méiose est une division cellulaire qui ne concerne que les cellules germinales (gamètes et cellules
souches). Elle aboutit à la naissance de quatre cellules (gamètes à n chromosomes). Il s’agit d’une
succession de deux divisions : la division réductionnelle et la division équationnelle.
- Individualisation des chromosomes.
- Disparition de l’enveloppe nucléaire.
- Appariement de chromosomes homologues
(2n) avec formation de chiasmas et
possibilité de crossing-over.
Rangement des paires de chromosomes
homologues de part et d’autre du plan
équatorial.
Séparation des chromosomes homologues
qui migrent aux pôles opposés de la cellule
(ascension polaire).
Première division cytoplasmique pour
donner deux cellules filles haploïdes avec un
exemplaire de chaque chromosome
possédant 2 chromatides (n2q).
Deuxième division cytoplasmique qui
permet l’obtention de 4 cellules filles
haploïdes dont les chromosomes n’ont plus
qu’une chromatide (nq).

b- La fécondation rétablit la diploïdie
La fécondation permet le rétablissement de l’état diploïde. C’est lors de celle-ci que les gamètes
subissent une duplication (leurs chromosomes, formés d’une seule chromatide, reconstituent à partir de
cette dernière la chromatide manquante : on est donc à n2q). Une fois leur matériel génétique dupliqué,
il y a fusion des noyaux (caryogamie) et l’on obtient une cellule à 2n2q.
B- La reproduction sexuée est à l’origine d’un brassage génétique
1- Chez les organismes haploïdes
Le brassage du matériel chromosomique chez un champignon comme Sordaria est révélé par la
disposition des spores jaunes et noires dans les asques. Les crossing-over et les positionnements
aléatoires des chromosomes et des chromatides expliquent les différents regroupements observés.
Formation d’asques de type 4/4 :
Formation d’asques de type 2/4/2 ou 2/2/2/2 :
2- Chez les organismes diploïdes
a- Le brassage intrachromosomique
Le crossing-over, qui s’effectue en prophase I, est responsable du brassage intrachromosomique. Il est
à l’origine des cellules haploïdes de type recombiné, c’est-à-dire qu’elles représentent de nouvelles
associations alléliques. S’il n’y a pas eu de recombinaison, les gamètes obtenus sont dits de type
parental.

Suite à leur appariement lors de la prophase I, les chromosomes homologues se séparent, sauf au
niveau de contacts particuliers, les chiasmas. Les chiasmas correspondent à des zones où se produisent
des échanges entre deux chromatides de chromosomes homologues.
Le brassage intrachromosomique :
b- Le brassage interchromosomique
Les paires de chromosomes homologues ont un comportement indépendant lors de leur séparation en
anaphase I. La répartition au hasard des chromosomes de part et d’autre du plan équatorial assure un
brassage interchromosomique lors de la première division de méiose.
Les chromosomes subissant ce brassage ont pu déjà être remaniés par des crossing-over. Dans ce cas, il
y a également brassage interchromosomique lors de l’anaphase II. Il y a donc une multiplication des
effets des deux types de brassages, augmentant ainsi la diversité génétique obtenue chez les nombreux
gamètes.
Le brassage interchromosomique :
c- Le rôle de la fécondation
Au cours de la fécondation, la rencontre d’un gamète mâle et d’un gamète femelle se fait au hasard.
Chaque gamète obtenu par méiose est donc susceptible de s’unir. Par conséquent, la fécondation
amplifie les deux types de brassage de la méiose.
Chez un organisme diploïde, les brassages génétiques associés à la fécondation conduisent à l’unicité
génétique des individus.
II- Innovations génétiques et évolution
A- L’origine du polymorphisme
1- La notion de polymorphisme
Tous les individus d’une espèce possèdent les mêmes gènes. Toutefois, il existe des variations
individuelles des gènes, les allèles. Ces versions du gène sont localisées au même endroit sur les

chromosomes (locus). L’ensemble des différents allèles d’un même gène au sein d’une population
correspond au polyallélisme.
Au sein d’une espèce, la fréquence des allèles est variable. On dit qu’un gène est polymorphe lorsqu’il
possède au moins deux de ses allèles en fréquence supérieure ou égale à 1%. Le polymorphisme
génique est donc une source de variabilité des individus qui, tout en ayant les mêmes gènes, ont une
combinaison allélique unique.
2- L’origine du pollyallélisme : les mutations
Une mutation est un changement dans la séquence nucléotidique de l’ADN. Il existe trois types de
mutations :
- La substitution qui correspond au remplacement ponctuel d’une base azotée par une autre.
- L’addition qui résulte de l’insertion d’un ou de plusieurs nucléotides dans la séquence du gène.
- La délétion ou perte d’un ou plusieurs nucléotides dans la séquence d’ADN.
Une substitution d’une base azotée peut être silencieuse du fait de la redondance du code génétique
c’est-à-dire qu’à un acide aminé peuvent correspondre plusieurs codons. Dans le cas d’un nouvel acide
aminé obtenu, la mutation est dite faux-sens. Si il y a obtention d’un codon stop, la mutation est alors
dite non-sens.
Une délétion ou une addition peuvent engendrer un décalage du cadre de lecture des nucléotides qui se
lisent par trois. La protéine verra alors un changement important des acides aminés la constituant.
Les trois types de mutations peuvent changer la structure et/ou le fonctionnement de la protéine. Leur
transmission à la descendance, ne s’effectuera que si les mutations sont faites au sein des cellules
germinales.
B- La création de nouveaux gènes
L’étude de différents gènes similaires a permis d’établir que certains gènes ont été obtenus par
duplication d’un gène sur le même chromosome ou sur deux chromosomes différents (transposition). Il
en résulte la création d’un nouveau gène puisque leur localisation est différente.
Pour un individu donné, il existe donc des familles multigéniques, ensemble de gènes obtenus à partir
d’un gène ancestral. Les deux gènes évoluent alors indépendamment l’un de l’autre. Ils peuvent subir
au cours de leur existence des mutations différentes ce qui engendre des divergences d’expression.
La duplication (avec ou sans transposition) et les mutations sont source d’innovations génétiques aux
conséquences phénotypiques variées. Celles-ci s’observent aussi bien au cours de l’évolution d’une
espèce que pour la création d’espèces nouvelles.
C- Mutations et avantage sélectif
La sélection naturelle opère sur des individus différents et sur des allèles différents. La fréquence
allélique des individus sélectionnés augmente, alors que celle des individus éliminés diminue.
Finalement, c’est la population qui évolue par des variations de fréquences alléliques. La sélection
naturelle, par le jeu de facteurs du milieu, trie les individus les plus aptes ; elle privilégie donc la
conservation des allèles ou associations d’allèles favorables aux conditions du moment.
Certaines mutations se font sans avantage sélectif : il s’agit de mutations neutres (ex. les hémoglobines
des mammifères).
 6
6
1
/
6
100%