elements de correction du ds n° 2

ELEMENTS DE CORRECTION DU DS N° 1 du 16/10/06
Intro 1
Conclusion 1
Schéma bilan 2
Thème 1 : 3
Thème 2 : 3
Thème 3 : 6
Thème 4 : 3
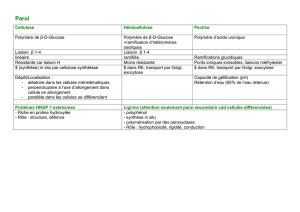
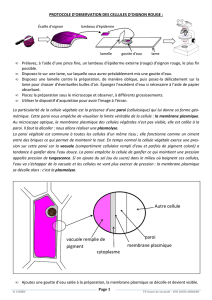
La paroi est une formation extracellulaire, une matrice fabriquée par la cellule végétale ; c’est
un édifice supramoléculaire essentiellement cellulosique.
Croquis possible d’une cellule végétale.
On peut s’interroger sur la mise en place des constituants de cette paroi, sur les propriétés
qu’ils lui confèrent mais aussi sur les contraintes qu’ils créent notamment vis à vis de la
croissance cellulaire.
Thème 1 :
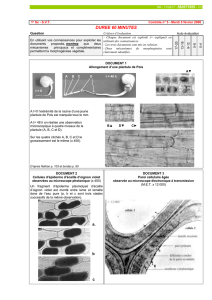
Document 1 : la cellulose représente 9% des constituants cellulaires au jour 5 pour en
constituer la presque totalité (94%) au jour 29.
L’enrichissement en cellulose, polymère du glucose, se fait au détriment des autres
constituants pariétaux : protéines, sucres neutres et acides uroniques sont quantité négligeable
au 29° jour.
La fibre de coton est ainsi une cellule dont la composition moléculaire pariétale est
complètement modifiée en 25 jours ; différenciée, la paroi est constituée essentiellement de
cellulose.
explication de la linéarité de la chaîne polyosidique.
Document 2 : il montre une cellule avec une forme allongée , étroite, éloignée de la forme
parallélépipédique de la cellule embryonnaire. La cellule est différenciée sur le plan
morphologique.
Document 3 : la coupe transversale de la fibre de coton révèle une section cylindrique avec
une paroi épaisse représentant les 3/4 du rayon et une lumière réduite dépourvue de contenu
cellulaire visible. La paroi est constituée de couches concentriques ; on peut supposer d’après
le document 1 que ce sont des dépôts successifs de cellulose (de fibres de cellulose).
La cellule fabrique la cellulose et l’assemble sur la face extracellulaire de la membrane
plasmique, grâce notamment à des enzymes.
Réalisation d’un schéma d’interprétation du doc3
Conclusion : dans la graine de coton, les cellules se différencient en cellules allongées
nommées fibres constituées essentiellement de cellulose située dans la paroi.
Comment la paroi intervient-elle dans la vie cellulaire et interfère-t-elle dans les relations
entre cellules, c’est à dire au niveau des tissus et organes ?
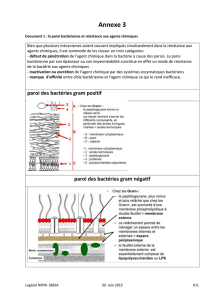
Thème 2

Les cellules observées sur le document 4a possèdent une paroi très épaisse (de 10 µm
environ) ; celle-ci est constituée de diverses couches concentriques claires et d’un « ciment »
sombre, liant les différentes cellules entre elles. Le détail de la paroi proposé par le document
4b présente les fibrilles de cellulose disposées avec des orientations définies qui changent
rythmiquement d’une strate à l’autre. On peut formuler l’hypothèse qu’il s’agit là d’une paroi
secondaire. Cette disposition des fibrilles augmente les propriétés de résistance des cellules et
permet la fonction de soutien du tissu.
Document 5a : les cellules épidermiques sont jointives, la vacuole turgescente plaque la
membrane plasmique contre la paroi.
Document 5b : après plasmolyse et traitement enzymatique, destiné à faire disparaître la paroi,
la cellule adopte une forme parfaitement sphérique. L’ablation de la paroi entraîne la perte de
la forme de la cellule et de la cohésion tissulaire.
Les protoplastes obtenus doivent être maintenus dans une solution isotonique pour ne pas être
lysés, la paroi confère un cadre rigide à la cellule, jouant le rôle d’un « exocytosquelette ».
Document 6 :
On constate que plus la branche est âgée, plus les microfibrilles de cellulose s’alignent dans
l’axe de celle-ci lorsque l’on s’éloigne du tronc assurant une plus grande rigidité de l’organe
et lui permettant de ne pas ployer sous son propre poids. En revanche la base de la branche
garde elle une certaine flexibilité, permise par la disposition transverse des fibres de cellulose.
La disposition des fibres de cellulose et l’épaisseur des dépôts jouent un rôle fondamental
pour conférer résistance, élasticité et flexibilité.
Les propriétés de résistance de la paroi végétale s’observent à tous les niveaux d’organisation
(cellule, tissu, organe) ; la paroi assure des fonctions de soutien indispensables en milieu
terrestre où la gravité est importante.
La cellule végétale est ainsi entourée par une matrice qui assure une cohésion avec les cellules
limitrophes, qui procure forme et cadre protecteur. Cependant, on peut s’interroger sur les
possibilités pour la cellule de déformation et de croissance dans ce contexte.
Thème 3
Document 7 :
L’auxine stimule la croissance de l’hypocotyle, d’un facteur 4 au bout de 2 heures,
l’augmentation est brutale mais après un temps de latence de 15-20mn environ ce qui suggère
l’existence de mécanismes complexes. Conjointement, en présence d’auxine, les échantillons
stimulés provoquent une diminution du pH environnant vraisemblablement due à une
sécrétion d’ions H+ par le cytoplasme vers le milieu extracellulaire. Le temps de latence plus
court (8-10mn) permet de formuler l’hypothèse que la diminution de pH précède l’élongation.
Document 8 :
L’expérience confirme le fait qu’un pH acide appliqué de manière exogène peut à lui seul
stimuler la croissance, la baisse de pH est donc la cause de l’élongation.
Le pH légèrement acide provoque une augmentation de la croissance importante et durable
alors qu’un pH très acide de 3 provoque une nette augmentation mais non durable ; on

observe même, au bout de 30 minutes, une croissance négative ce qui traduit l’effet létal
(mortel) du pH trop bas.
Document 9 :
Un pH légèrement acide ne provoque pas cette fois ci d’extension pariétale alors qu’un pH
acide de 3 provoque une stimulation de l’extensibilité pariétale durable.
La confrontation des deux documents permet de conclure qu’il existe une nette différence
entre la stimulation de la croissance et celle de l’extensibilité pariétale. Une simple baisse de
pH n'agit donc pas directement (effet physico-chimique) puisque l'effet d'un pH très acide agit
sur les propriétés de la paroi isolée mais pas sur l'échantillon vivant. Au contraire, une baisse
de pH, dans un registre physiologique, permet une stimulation importante de la croissance des
échantillons vivants mais n'a pas d'effet simplement physico-chimique sur les parois isolées.
On conclut de cette série d'expériences, que la régulation de la croissance par la baisse de pH
induite par l'auxine nécessite un processus biologique et pas seulement physico-chimique.
Document 10 :
D’après le modèle proposé, les hémicelluloses sont liées aux molécules de cellulose grâce à
des liaisons faibles ; ces liaisons sont dépendantes du pH du milieu environnant ; on peut
formuler l’hypothèse qu’une acidification du milieu rompt les liaisons H autorisant la
séparation des différents constituants nécessaire pour la croissance pariétale . Les expansines
pourraient jouer un rôle dans le contrôle de cette croissance.
Reste à identifier le moteur de la croissance…
Document 11 :
- pour les régions (b)
La déformation est positive ou négative selon la concentration du milieu. Lorsque la
déformation est positive, elle ne se stabilise pas et s'accroît au contraire pendant plusieurs
heures. Lorsque les échantillons sont replacés dans de l'eau pure, ils n'atteignent pas la taille
de celui qui était dans l'eau depuis le début de l'expérience mais reprennent tous une
croissance à vitesse égale.
Le mannitol ne pénétrant pas dans les cellules, il est responsable de la pression osmotique du
milieu extracellulaire. Le milieu contenant 0.4M de mannitol est un milieu hypertonique,
l’eau est sortie de la cellule diminuant ainsi la pression de turgescence. En replaçant les
échantillons dans de l’eau pure, on provoque l’entrée d’eau dans les cellules d’où
l’augmentation de la pression de turgescence, moteur de l’élongation cellulaire.
- pour les régions (f)
Chez un échantillon en fin de croissance, les déformations observées sont réversibles et
dépendent essentiellement des entrées et des sorties d'eau dues à la turgescence relative des
cellules, provoquée par le potentiel hydrique du milieu.
La paroi secondaire déjà présente bloque toute possibilité d’élongation.
En conclusion, la croissance de la cellule végétale ne peut se réaliser que grâce à des
changements des propriétés de la paroi primaire ; l’auxine, hormone végétale, modifie le pH
extracellulaire ce qui entraîne une réorganisation des principaux constituants pariétaux sous le

contrôle des expansines. La pression de turgescence est le principal moteur de l’élongation
cellulaire.
Les rôles des parois primaire et secondaire, essentiellement cellulosiques et des protéines
associées ont été mis en évidence. Qu’en est-il pour la lamelle moyenne ?
Thème 4.
Document 14 :
La lamelle moyenne des cellules de pulpe de fruit charnu est très épaisse (50 fois plus que
celle d’une cellule de parenchyme). Cette lamelle est composée de pectines c'est-à-dire de
composés de nature gélifiantes chargés grâce à la présence de fonctions acides : polyuronates.
Les ions Ca++ réalisent des ponts entre les différents polymères. La consistance des tissus
charnus et leur richesse en eau peut en partie s’expliquer par l’épaisseur de la lamelle
moyenne.
Document 12 : Le concombre est un fruit charnu, d’après le document précédent, on peut
s’attendre à ce que la lamelle moyenne soit épaisse et soit impliquée dans les processus
auxquels les documents font référence.
Une comparaison par lot permet de ne s’intéresser qu’à la variation d’un seul paramètre à la
fois.
Lot 1 : le calcium intracellulaire ne varie pas alors que le calcium pariétal augmente
linéairement quand on s’éloigne du puits. Hypothèse : la blessure entraîne une remobilisation
du calcium pariétal au niveau de la blessure.
Lot 2 : même remarque que précédemment pour le calcium intracellulaire mais augmentation
plus importante du calcium pariétal. Hypothèse : le végétal réagit à l’infection par le
champignon (absent dans le lot 1) en mobilisant le calcium pariétal encore plus.
Lot 3 et lot 4 : la quantité de calcium intégré à la paroi, donc lié aux pectines, augmente avec
la quantité de calcium ajouté ; on assiste en parallèle à une diminution de l’infection.
Hypothèse : le calcium pariétal limite l’infection parasitaire.
Conclusion : la lamelle moyenne joue un rôle protecteur vis à vis des infections mycéliennes.
Dans le thème 2, son rôle dans l’adhésion intercellulaire a été mis en évidence.
Document 13 : On constate une diminution de la concentration du milieu en acide
galacturonique avec l’augmentation de la concentration en calcium. Or l’acide galacturonique
est le produit de l’action de l’enzyme sur les pectines. Si, il y a moins de produits formés,
c’est que l’activité enzymatique est plus réduite en présence de concentrations élevées de
calcium. e calcium pariétal inhibe donc la polygalacturonase du champignon.
La paroi et plus spécifiquement ici la lamelle moyenne joue un rôle dans la protection de
l’organisme en limitant l’infection du parasite fongique.

Thème 5 : Schéma bilan
Organisation fonctionnelle de la paroi des cellules végétales
1
/
5
100%