Polymorphisme génétique : Importance et maintien

Le polymorphisme génétique et son maintien
Introduction :
La diversité phénotypique est une des propriétés générales des populations naturelles.
Dans celles-ci, les individus présentent pour la plupart des caractères très différents. Une part
de cette variabilité résulte de différences entre génotypes, d’où une diversité génétique sous-
jacente.
Or, elle est, à l’heure actuelle, source de préoccupation car on constate un phénomène
d’érosion de la biodiversité suite à l’activité humaine. Dans les populations naturelles, cette
variabilité génétique se manifeste par le polymorphisme génétique, c’est à dire le nombre de
loci polymorphes (loci présentant de multiples allèles différents).
On peut alors se demander pourquoi le polymorphisme, et donc cette diversité, revêt une
telle importance.
Pour répondre à ce problème, on va donc essayer, dans un premier temps, d’expliquer
l’importance évolutive du polymorphisme, puis d’étudier comment il peut être maintenu.
I. Importance du polymorphisme
1. Mesure du polymorphisme
Pour mesurer le polymorphisme, on peut utiliser deux méthodes : celles employant les
allozymes et celles utilisant les fragments de restriction.
Le polymorphisme des allozymes
Ces enzymes se différencient par leur mobilité électrophorètique et elles correspondent à
des allèles différents à un même locus. La variabilité des allozymes dans une population est
donc un indicateur de la variabilité génétique. On a trouvé une importante diversité parmi les
allozymes dans presque toutes les populations naturelles.
Le polymorphisme des allozymes est largement répandu chez les organismes supérieurs.
Mais bien que le polymorphisme génétique soit largement répandu, il n’est pas universel.
Par exemple, deux sous-espèces de guépard (Acinomynx jubatus) sont pratiquement
monomorphes.
Apparemment, la population de guépard, dont l’aire de répartition était mondiale à une
certaine époque, a subi au moins deux diminutions importantes du nombre des individus ce
qui a provoqué la perte d’une grande part de leur variabilité génétique (O’Brien et al. 1987).
Document 9: Exemple de gel montrant un polymorphisme d’alllozymes
Le polymorphisme des fragments de restriction de l’ADN
La source fondamentale de la variabilité génétique se situe au niveau des séquences d’ADN.
On peut révéler les polymorphismes des séquences d’ADN à des sites précis de la séquence
de nucléotides (sites de restrictions).
Les polymorphismes qu’entraînent la présence ou l’absence d’un site de restriction, sont
reconnaissables parce que la longueur des fragments de restriction est modifiée. Ces
polymorphismes des fragments de restrictions sont en fait, des polymorphismes de la
longueur des fragments de restrictions (RFLP). Ces RFLP sont répartis à travers le génome
et sont répandus chez presque tous les organismes. Les RFLP ont une grande importance en
génétique humaine comme marqueurs génétiques pour cartographier le génome et comme
marqueurs génétiques de maladie. On peut donc estimer le degré de polymorphisme au niveau
nucléotidiques à partir de données sur les sites de restrictions.

Utilisation du logiciel ANAGENE pour montrer le polymorphisme des fragments de
restriction
De même, pour comparer des gènes différents et des populations différentes, il est
nécessaire d’avoir une mesure quantitative et pratique de la variabilité génétique. On peut
quantifier la variabilité génétique en utilisant le concept de fréquence allélique.
Dans un groupe d’individu, la fréquence allélique d’un allèle donné est simplement la
proportion au locus considéré de tous les allèles de ce type. Pour un gène considéré
comprenant 2 allèles A et a, cette fréquence dans un échantillon est donc égale :
Chaque homozygote chacun porte
porte 2 copies de cet allèle deux allèles à ce locus
f(A) = [ 2*n(AA) + n(Aa)] / 2*N = [n(AA) + 0.5 n(Aa)] / N
chaque hétérozygote
porte une copie
D’où f(A)= f(AA) + 0.5 f(Aa)
où n (AA) = nombre de génotypes homozygotes pour cet allèle,
n (Aa) = le nombre de génotypes hétérozygotes comportant ces allèles
N = nombre d’individus totaux
f (AA) = fréquence du génotype AA
Cette estimation de la fréquence allélique est proche de la vraie valeur de la fréquence dans
la population si l’échantillon est suffisamment grand.
Le concept de fréquence allélique sous-tend l’idée de polymorphisme.
Maintenant, que nous avons vu comment on peut estimer le polymorphisme dans les
populations, nous allons essayer de comprendre quel avantage il peut y avoir à être
polymorphes ?
2. Rôle du polymorphisme dans un environnement changeant
L’environnement, dans lequel vivent les populations naturelles, n’est pas stable au cours du
temps (notion de stochasticité environnementale). De nombreux facteurs fluctuent comme la
température, la luminosité, les ressources alimentaires ou encore les relations interspécifiques.
Certaines de ces modifications sont périodiques donc les êtres vivants y sont généralement
habitués. Il existe cependant des facteurs apériodiques qui sont aléatoires à l’échelle de la
micro-évolution ; c’est le cas notamment de bon nombre de changements dus à l’homme.
Pour que les organismes puissent y faire face, ils doivent s’adapter. Or, si dans une
population donnée, tous les individus présentent le même génotype et que le milieu change
soudainement, la population est vouée à l’extinction. Par contre, si certains individus ont des
allèles autres qui présentent un avantage dans certaines conditions environnementales, ils
seront sélectionnés (survie meilleure, taux de reproduction plus importante) et la population
pourra ainsi se maintenir.
Illustrons ceci par un exemple célèbre : celui du mélanisme industriel chez la Phalène du
bouleau (Biston betularia).
Document 6: Camouflage des deux morphes de la Phalène du bouleau (Biston betularia)
sur un tronc couvert de lichens

En Europe du nord, cette espèce de papillon présente une coloration crème clair, mouchetée
de marron. Lorsque pendant la journée, ce papillon de nuit se pose sur une branche, sa couleur
le camoufle à la vue de certains prédateurs (= cryptisme). Ceci est vrai, si cette branche est de
couleur claire, ce qui est généralement dû à sa couverture de lichens.
Au cours de la révolution industrielle, en Grande Bretagne, la pollution par la fumée a
localement tué les lichens, ce qui a noirci les troncs sur lesquels se posaient les phalènes. On a
constaté après coup que dans les collections datant de cette époque (vers 1830) une forme
mélanique de ce papillon, bien camouflée dans les branches sombres, est devenue de plus en
plus abondante. Au cours du 19ème siècle, l’importance de la forme mélanique a augmenté
dans les régions industrielles, au point que cette forme est devenue la plus fréquente.
Cet accroissement de fréquence a été presque certainement dû à la sélection naturelle. En
effet, les observations de Kettlewell, au 20ème siècle, ont montré que les oiseaux mangent les
papillons mal camouflés. D’ailleurs, si on relâche dans une région industrielle un mélange de
2 formes de papillons, ce sont les individus mélaniques qu’on a le plus de chances de
recapturer, vivants, plus tard. Au contraire, si c’est dans une région non industrielle qu’on
effectue la même expérience, la plupart des papillons recapturés sont clairs.
A noter ici que dans la plupart des cas de mélanisme industriel, le motif de coloration noire
est déterminé par un seul allèle dominant.
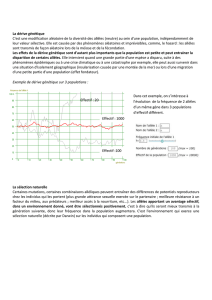
Comme le montre cet exemple, la sélection naturelle va faire fluctuer les fréquences
alléliques en fonction des avantages que procurent tels ou tels allèles. Le polymorphisme
génétique est donc très important dans un environnement changeant.
De même, il existe une relation positive entre le polymorphisme génétique et le degré
d’hétérozygotie. En effet, plus la proportion de gènes polymorphes est élevée dans une
population, plus nombreux sont les gènes que l’on peut retrouver sous la forme hétérozygote
chez un individu quelconque. Or, l’hétérozygotie revêt une grande importance pour les
populations naturelles car elle est souvent associée à la vigueur hybride appelée aussi
l’hétérosis.
3. Rôle de la vigueur hybride
L’hétérozygotie correspond au nombre de loci hétérozygotes qui existent dans une
population donnée. Ainsi, il permet, tout comme le polymorphisme, de quantifier la diversité
génétique d’une population.
Exemple de croisement entre deux individus de race pure :
Individu A de Individu B de
génotype AA génotype aa
Individu C hybride de génotype Aa
L’individu C dans un environnement donné a une viabilité plus grande.
Tout d’abord, il présente deux allèles différents donc il (ou plutôt ses descendants) pourra
faire face aux variations environnementales plus facilement que ces parents. Il sera ainsi
sélectionné (taux de survie et taux de reproduction plus important).
De plus, l’hétérozygotie permet de camoufler et d’empêcher l’expression d’allèles délétères
récessifs. Ainsi, si l’allèle a induit un disfonctionnement, l’individu B de race pure sera donc
malade alors que l’individu C (si on considère l’allèle A dominant) ne le sera pas. C’est ce
que l’on observe dans le cas de l’anémie falciforme. Cet exemple sera traité plus en détail
dans la dernière partie.
Par contre, l’individu C pourra transmettre la maladie à sa descendance comme le montre le
tableau de croisement ci-dessous.

Allèles des parents
HbβA
HbβS
HbβA
HbβA/HbβA
HbβS/ HbβA
HbβS
HbβS/ HbβA
HbβS/ HbβS
Remarque : HbβA : l’allèle normal ; HbβS : l’allèle mutant.
Tableau de croisement entre deux individus hétérozygotes (HbβS/ HbβA)
¼ des individus ainsi obtenus sont homozygotes (HbβA/HbβA) pour cet allèle ; ils ont une
hémoglobine normale. La moitié des individus sont hétérozygotes comme leurs parents et ¼
sont homozygotes (HbβS/ HbβS) : ils sont donc profondément anémiés.
Nous avons montré ici l’importante de la diversité génétique et donc du polymorphisme
dans l’adaptation des populations au milieu. Mais pour maintenir le polymorphisme génétique
dans une population il faut que les fréquences alléliques restent approximativement constantes
au cours du temps ; c’est pourquoi nous allons nous intéresser aux différentes forces
évolutives qui peuvent influer sur ces fréquences.
II. Polymorphisme et les forces évolutives
1. Loi d’Hardy-Weinberg
Afin d’estimer les variations des fréquences alléliques et par-là même les fréquences
génotypiques, nous utilisons des modèles mathématiques. Un modèle de prédiction des
fréquences génotypiques utilisé en génétique des populations est celui d’Hardy-Weinberg.
Ce modèle sous-tend plusieurs hypothèses :
a. L’organisme considéré est diploïde.
b. La reproduction est sexuée.
c. Les générations sont non chevauchantes. Cela correspond à un cycle de reproduction
très simple dans lequel les individus de chaque génération meurent avant la naissance des
membres de la génération suivante (ex : plante annuelle). En plus, les générations sont séparés
dans le temps (génération discrète).
d. Les croisements se font au hasard (panmixie).
e. La taille des populations est très grande.
f. Les migrations sont négligeables.
g. On peut ignorer les mutations.
h. La sélection naturelle n’a pas d’effet sur ces populations naturelles.
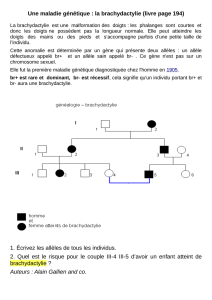
Prenons un exemple : on va considérer un locus à deux allèles A et a, de fréquences
alléliques égales respectivement à p = f(A) et q = f(a) (où p+q = 1). Deux individus de la
génération n vont se croiser au hasard d’où l’union au hasard des gamètes (Hypothèse d), à la
génération suivante n+1, on aura :
Gamètes des parents
A (p)
a (q)
A (p)
AA (p2)
Aa (pq)
a (q)
Aa (pq)
aa (q2)
f(AA)n+1 = p2 ; f(Aa) n+1 = 2pq et f(aa) n+1 = q2
Les fréquences alléliques à la génération suivante seront donc égales à :

p n+1 = f(AA) + 0.5* f(Aa) = p2 + 0.5*2pq = p2 + pq = p[p+q] = p
p n+1 = p
q n+1 = f(aa) + 0.5* f(Aa) = q2 + 0.5*2pq = q2 + pq = q[p+q] = q
q n+1 = q
Les fréquences alléliques à la nouvelle génération sont donc exactement les mêmes qu’à la
génération précédente. Lorsque les croisements se font au hasard, les fréquences alléliques
restent donc constantes. A toutes les générations, par conséquent, les fréquences génotypiques
sont : p2, 2pq et q2 pour AA, Aa et aa respectivement, et ces fréquences constituent ce qui est
souvent appelé l’équilibre d’Hardy-Weinberg.
La constance des fréquences alléliques (et de ce fait, celle de la composition génotypique de
la population) est la conséquence la plus importante de la loi d’Hardy-Weinberg. Cette
constance implique, en absence de forces évolutives particulières qui modifieraient les
fréquences alléliques, la conservation de la variabilité génétique telle qu’elle est et donc du
polymorphisme.
Document 10 : Exercice pour tester l’hypothèse de panmixie
C’est un modèle donc une simplification de la réalité. Une telle population n’existe pas, ceci
permet simplement de faire des prédictions.
Nous allons voir maintenant les facteurs qui peuvent modifier cet équilibre, en commençant
par la consanguinité et de la dérive.
2. Rôle de la consanguinité et de la dérive
Il existe plusieurs modes de reproduction ; dans le cadre du modèle d’Hardy-Weinberg, on a
considéré une reproduction sexuée avec des croisements panmictiques (Hypothèse b et d). On
va s’intéresser à un autre mode de fécondation : l’autofécondation (= autocroisement).
Dans le cas d’autofécondation, les fréquences alléliques vont évoluer ainsi :
Génotype considéré
AA
Aa
aa
Fréquence
génotypique à la
génération (n)
F(AA)n
F(Aa)n
F(aa)n
Génotype obtenu à
la génération (n+1)
AA
¼ AA, ½ Aa, ¼
aa
aa
Fréquence
génotypique à la
génération (n+1)
F(AA)n+1
F(Aa)n+1
F(aa)n+1
F(AA)n+1 = F(AA)n + ¼ F(Aa)n+1
F(Aa)n+1 = ½ F(Aa)n+1
F(aa)n+1 = F(aa)n + ¼ F(Aa)n+1
Remarque : p est toujours égale à la fréquence de l’allèle A et q de a.
Comme on peut le voir ici, à chaque fécondation le nombre d’hétérozygotes diminue de
moitié à chaque génération d’où une perte de la diversité génétique, qui peut se traduire à long
terme par la fixation d’un des allèles, c’est à dire la perte de l’autre allèle ( (F(AA) =1 et F(aa)
=0)).
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%