La procréation

1
Récapitulatif de l’ensemble des
chapitres de l’année.
L’an dernier vous avez pu constater avec l’épreuve de français qu’il n’est pas facile de se souvenir des chapitres vus
en début d’année. On a tendance à croire qu’on ne se souvient de rien ! La peur du trou de mémoire est anxiogène.
Or cette année, c’est dans toutes les matières que votre mémoire va devoir être performante ! D’où l’idée de vous
proposer un fichier récapitulatif de l’ensemble des chapitres de l’année.
Ce fichier qui vous est proposé est une trame de tous les chapitres que nous allons aborder. Vous y trouverez :
-la période à laquelle nous allons les travailler ;
-les parties du Bulletin Officiel (B.0) écrit par l’éducation nationale ;
-les introductions avec les problématiques ;
-les plans des TP du livre ;
-les plans du cours avec quelques notions ;
-il y aura aussi quelques définitions de mots compliqués.
En théorie, vous pourriez vous présenter à l’épreuve du BAC en connaissant par cœur les parties du B.O. En
pratique, notre travail cette année va consister à expliquer les notions du B.O notamment avec les documents du
livre. Aussi, plus cette année qu’une autre, votre livre constitue votre référence. Si vous êtes absent, si vous vous
n’avez pas compris une idée, si vous avez oublié quelque chose, vous devriez être en mesure de vous en sortir en
lisant votre livre.
De plus, votre mémoire fonctionne sur la répétition, vous le savez bien, mais aussi sur la classification : chaque
chose trouve une place dans votre cerveau comme l’arborescence d’un disque dur. Or, il n’existe pas de meilleur
rangement qu’un livre ! Plus vous apprendrez ce qu’il y a dans votre livre, plus votre mémoire sera performante !
Bonne année à tous.
Michel Patalano.

2
Sommaire :
Partie 1 : Parenté entre êtres vivants actuels et fossiles. Phylogenèse.
4 semaines=>De septembre à début octobre
Chapitre 1 : Les relations de parentés entre les êtres vivants (p.3)
Chapitre 2 : La lignée humaine (p.5)
Partie 2 : Stabilité et variabilité des génomes. Evolution.
6 semaines=>De mi-octobre à mi-décembre
Chapitre 3 : Les innovations génétiques (p.7)
Chapitre 4 : Méiose, fécondation et stabilité de l’espèce (p.9)
Chapitre 5 : Méiose, fécondation et brassage génétique (p.11)
Chapitre 6 : Evolution et génétique (p.13)
Partie 3 : La mesure du temps dans l’histoire de la Terre et la vie
2 semaines=>Début décembre à fin décembre
Chapitre 7 : La mesure du temps (p.15)
Partie 4 : La convergence lithosphérique et ses effets
4 semaines=>Début janvier à début février
Chapitre 8 : Convergence et subduction (p.17)
Chapitre 9 : convergence et collision continentale (p.19)
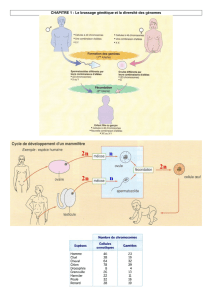
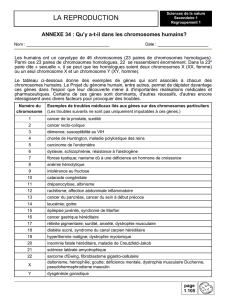
Partie 5 : La procréation
6 semaines=>De mi-février à fin mars
Chapitre 1O : Du sexe génétique au sexe phénotypique (p.21)
Chapitre 11 : régulation physiologique de l’axe gonadotrope (p.23)
Chapitre 12 : Comportement sexuel, maîtrise de la reproduction (p.25)
Partie 6 : La défense immunitaire
4 semaines=>De début avril à fin avril
Chapitre 13 : SIDA et système immunitaire (p.27)
Chapitre 14 : les processus immunitaires (p.29)
Partie 7 : Le couplage des évènements biologiques et géologiques au cours du temps
1,5 semaine=>Début mai à mi-mai.
Chapitre 15 : Crises biologiques et histoire de la Terre (p.31)

3
Partie 1 : Parenté entre êtres vivants actuels et fossiles. Phylogenèse.
4 semaines=>De septembre à début octobre
Chapitre 1 : Les relations de parentés entre les êtres vivants
B.O :
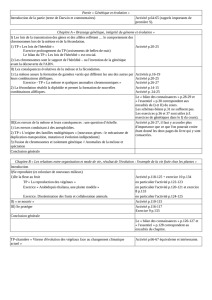
L’établissement de relations de parenté entre les vertébrés actuels s’effectue par comparaison de caractères
homologues : embryonnaires, morphologiques, anatomiques et moléculaires.
Les comparaisons macroscopiques prennent en compte l’état ancestral et l’état dérivé des caractères.
Seul le partage d’états dérivés des caractères témoigne d’une étroite parenté.
Ces relations de parenté contribuent à construire des arbres phylogénétiques.
Les ancêtres communs représentés sur les arbres phylogénétiques sont hypothétiques, définis par l’ensemble des
caractères dérivés partagés par des espèces qui leur sont postérieures.
Ils ne correspondent pas à des espèces fossiles précises.
Une espèce fossile ne peut être considérée comme la forme ancestrale à partir de laquelle se sont différenciées
les espèces postérieures.
Introduction
Matériel : grenouille, souris, poisson, reptile, tortue, lézard, salamandre
+ Un ornithorynque
Dans quel groupe va-t-on classer l’Ornithorynque ?
Diversité incroyable d’animaux comme de plantes
Une organisation commune qui reflète les liens de parenté… donc on peut construire des arbres généalogiques sans
ancêtre identifié !
= arbre phylogénétique
Comment peut-on établir la généalogie d’animaux aussi divers qu’une truite, une grenouille, un lézard, un
pigeon ou encore un Gorille ?
Plan des TP du livre (page 28)
TP1 : Recherche de parenté grâce à des arguments embryologique (p.20)
TP2 : Recherche de parenté grâce à des arguments anatomiques (P.22)
TP3 : Recherche de parenté grâce à des arguments moléculaires (P.24)
TP4 : L’utilisation des arbres phylogénétiques (P.26)
Evaluation des Capacités Expérimentales
1-ECE : Globule rouge
2-ECE : Liens de parenté
Plan du cours :
1-Historique de la classification du vivant
1.1-De l’intérêt de classer les plantes
Utilisation médicale
1.2-Naissance de la taxonomie
- division des organismes en critères précis ;
- réunion en critères de similarité (Linné) ;
- règne : des vertébrés ;
- classe : des mammifères ;
- ordre : des Anthropoïdes ;
- famille : hominidés ;
- genre : Homo ;
- espèce : sapiens (voir p.18) ;
1.3-De la classification à la transformation des êtres vivants
Jussieu : caractères non équivalents
Choisir un petit nombre de caractères
Définir des taxons

4
Cuvier : application de cette classification aux animaux
Trouver du sens à la classification :
- Lamarck : théorie transformiste
(=hérédité des caractères acquis)
« Pourquoi les êtres vivants sont-ils différents ? »
- Darwin : « Pourquoi, malgré leur potentiel de variation, les êtres vivants se ressemblent-ils encore ? »
Début de la recherche de liens de parentés entre les EV
2-Le principe de l’établissement de parentés
Par comparaison de caractères homologues
= similitude de position c’est à dire une même origine embryologique
2.1-Etablissement d’arbres phylogénétiques à partir de données macroscopiques
Recherche de caractères « évolués » partagés
Arbre phylogénétique
=arbre généalogique sans ancêtre identifié
Parenté éloignée si partage d’un caractère primitif
Parenté étroite si partage d’un caractère dérivé
Tout caractère présente un état dérivé et un état ancestral
Groupe monophylétique :
Propriété d’un groupe comprenant l’ancêtre commun et ses descendants
(ex : les batraciens avec les anoures et les urodèles)
(contre-ex : le groupe des Gravigrades, proboscidiens et siréniens)
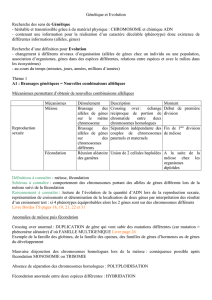
2.2-Etablissement de relations de parentés à partir de données moléculaires
2 molécules (Gène antennapaedia/Hox B6 ou Protéine Rhodopsine) :
Si similitude de leurs séquences d’acides aminés ou de nucléotides :
= molécules homologues (voir pages 24/25).
Degré de similitude : fruit de l’évolution
Peu de différences=espèces proches
Beaucoup de différences= espèces éloignées
3-Vitesse d’évolution moléculaire et macroscopique
3.1-Horloge moléculaire
Nombre de différences : proportionnel au temps écoulé
Vitesse de modification d’AA ou de nucléotides constante
=> horloge
(Voir page 30 le cas de l’hémoglobine)
3.2-Horloge paléontologique
Datation absolue des fossiles
estimation de la vitesse d’évolution macroscopique
Passage molaire à tubercule => molaire à lame : 50 Ma
100 Ma : grandes étapes évolutives
- Plan d’organisation des vertébrés : 500 Ma ;
- Membre tétrapode : 400 Ma ;
- œuf d’amniote : 300 Ma ;
- Mammifère : 200 Ma)
Documents page 31 à savoir par cœur.
Vocabulaire :
Etat dérivé d’un caractère :
Etat d’un caractère que partagent tous les individus formant un groupe issu d’un même ancêtre. L’ancêtre
hypothétique est celui qui a possédé le premier le caractère dans cet état
Polyphylétique :
Pas d’ancêtres communs, comme c’est le cas des poissons.
Paraphylétique :
Groupe des reptiles qui ont le même ancêtre mais dont un groupe est issu mais n’en fait pas parti comme les
oiseaux.

5
Chapitre 2 : La lignée humaine
B.O :
La place de l’homme dans le règne animal
L’homme est un eucaryote, un vertébré, un tétrapode, un amniote, un mammifère, un primate, un hominoïde, un
hominidé, un Homininé : ces caractères sont apparus successivement à différentes périodes de la vie.
L’homme partage un ancêtre commun récent avec le Chimpanzé et le Gorille.
Cet ancêtre commun n’est ni un Chimpanzé (ou un Gorille) ni un Homme.
La divergence de la lignée des Chimpanzé et de la ligné humaine peut-être située il y a 7 à 10 Ma.
Les critères d’appartenance à la lignée humaine
Les critères d’appartenance à la lignée humaine sont les caractères liés à la station bipède, au développement du
volume crânien, à la régression de la face et aux traces fossiles d’une activité culturelle.
On admet que tout fossile présentant au moins un de ces caractères dérivés appartient à la lignée humaine.
Le caractère buissonnant de la lignée humaine
La lignée humaine est représentée actuellement par une seule espèce.
Plusieurs espèces d’hominidés ont vécu entre 6 Ma et 100000 ans, époque où apparaissent les Homo sapiens.
Ces espèces appartiennent à deux genres : les Australopithèques et les Homo.
Les Australopithèques possèdent des caractères dérivés de la lignée humaine en rapport avec la bipédie.
Les espèces du genre Homo possèdent en outre des caractères dérivés crâniens marqués notamment par une
augmentation du volume crânien et une réduction de la face.
Les Australopithèques ont vécu entre 4Ma (A.anamensis) et 1 Ma (A.robustus.
Les Homo les plus anciens (H.habilis) sont datés de 2,5 Ma.
Plusieurs espèces d’homininés ont vécu en même temps.
Les Australopithèques formeraient un rameau de la lignée humaine détaché assez tôt de celui des homo.
Les espèces fossiles actuellement datées entre 4 Ma et 1,5 Ma sont toutes africaines. Cela peut s’expliquer par
l’origine africaine de la lignée humaine ou par les conditions de fossilisation exceptionnelles de la vallée du rift
africain.
Les Homo erectus sont connus d’abord en Afrique (adolescent du Turkana : 1,6 Ma) ; ils forment un groupe très
diversifié dont l’évolution est marquée notamment par l’augmentation graduelle du volume crânien.
De nombreuses populations colonisent l’Afrique du nord, l’Afrique du sud, le Proche Orient, l’Asie et l’Europe.
L’homme de Neandertal trouvé en Europe semble provenir de l’évolution d’Homo erectus ayant colonisé l’Europe.
L’origine des Hommes modernes
Toutes les populations actuelles partagent les mêmes allèles, avec une fréquence variable.
La population ancestrale n’aurait compté que quelques dizaines de milliers d’individus.
Homo sapiens serait une nouvelle espèce apparue en Afrique ou au Proche Orient il y a 100000 à 200000 ans et
aurait colonisé tous les continents en remplaçant Homo erectus.
Introduction :
L’homme : Piédestal
Coupé de la nature/Image de Dieu
Darwin : Conteste l’idée que les espèces animales soient immuables
En explique la cause : la Sélection naturelle
L’homme occupe une place à part
L’homme est le seul représentant de son espèce
Imaginez : Si nous pouvions croiser Neandertal dans la rue…
Pb : quelle est l’origine de cette espèce ?
Plan de la leçon :
1-La place de l’homme dans le règne animal
2-Les critères d’appartenance à la lignée humaine
3-Le caractère buissonnant de la lignée humaine
3.1-Les Australopithèques, Homininés les plus anciens :
3.2-Les premiers hommes en Afrique
4-L’origine des Hommes modernes
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%