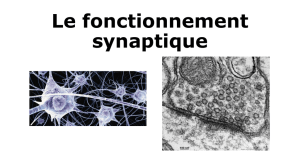
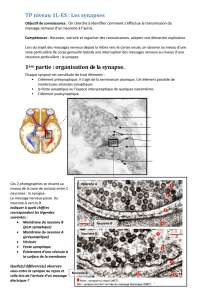
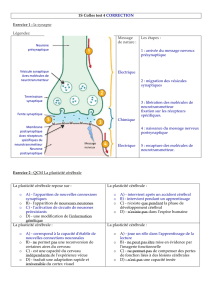
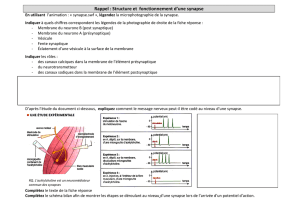
La mémoire à long terme

1
CM 2 du 08/11/01
Cours de physio
N
NE
EU
UR
RO
OB
BI
IO
OL
LO
OG
GI
IE
E
D
DE
E
L
LA
A
M
ME
EM
MO
OI
IR
RE
E
E
ET
T
D
DE
ES
S
A
AP
PP
PR
RE
EN
NT
TI
IS
SS
SA
AG
GE
ES
S.
.
Dans les 3 études des mécanismes impliqués dans la mémoire on constate 3 types
de résultats :
Observation de patients cérébrolésés (trouble de la mémoire et des
apprentissages). Le but est d’établir des liens entre la fonction atteinte
(tests psychologiques) et la structure lésée (analyses post mortem puis
scanner ou imagerie médicale).
Imagerie médicale sur des personnes saines pour observer les zones
cérébrales activées au cours d’une tache donnée. Liens entre les zones du
cerveau activées et la nature de la tache.
Limites : les techniques d’imagerie médicale.
Les techniques liées aux tests… il faut trouver des taches qui n’utilisent qu’une
zone précise de la mémoire.
Expérimentation animale.
Limites : le rat n’est pas un homme.
I. CLASSIFICATION
a) Les mémoires
1) Mémoire à court terme (MCT) et mémoire à long terme (MLT)
Cf. poly page 1.
Un système de mémoire nécessite, pour fonctionner, des capteurs qui permettent
l’entrée des données. Ils vont transmettre les données à un système de stockage. On va

2
pouvoir lire telle ou telle donnée à la demande. S’il y a un disfonctionnement à un niveau,
c’est tout le système qui sera atteint.
Le registre sensoriel.
Dans ce modèle il est une petite forme de mémoire car l’information est capable de
décider dans ce registre pendant une centaine de millisecondes.
La mémoire à court terme
Encodage. Reçoit des info du registre sensoriel mais aussi de la mémoire à long terme
(rappel). Les info sont maintenues directement accessibles pendant une vingtaine de
secondes.
La mémoire à long terme
Consolidation. Reçoit les info de la mémoire à court terme mais aussi du registre
sensoriel. Stockage définitif mais pas directement accessible, pour cela il faut un rappel
dans la MCT.
2) Mémoire à long terme déclarative et procédurale
Cf. doc. 3.
On a observé des patients amnésiques et des animaux.
Mémoire déclarative (explicite)
Capacité de garder en mémoire des évènements, info dont le point commun est qu’elles
peuvent être déclarées. Elles sont accessibles à la conscience, mémoire consciente qui
s’exprime par le langage ou par des images mentales.
Mémoire épisodique
Mémoire des évènements.
Mémoire sémantique
Mémoire du sens des choses.

3
Mémoire non déclarative (implicite)
C’est tout le reste.
Mémoire procédurale
Capacité d’apprendre des manières de faire sans forcément se souvenir comment on a
appris. Permet l’acquisition progressive d’habiletés (motrices, perceptivo-motrices,
cognitives) grâce à l’entraînement.
Amorçage
Facilitation de la détection d’un signal, du traitement d’une info à partir d’une
expérience récente.
Conditionnement classique
Apprentissage non associatif
b) Les apprentissages
1) Méthodes d’étude
Elles reposent sur l’utilisation de tests le plus spécifique possible d’un type
d’apprentissage (c’est difficile). On a :
Des apprentissages moteurs et visio-moteur
ex : test d’opposition des doigts de la main.
Test du rotor.
Des apprentissages visuels
ex : lecture des mots à l’envers.
discrimination d’une cible (cf. doc. 1).
Des apprentissages cognitifs
ex : test de résolution de problème.

4
Quel que soit le type de tache, on quantifie toujours l’apprentissage par
l’évolution de la performance au test. Cette performance évolue avec le nombre d’essais
puis stagne.
b) Les formes d’apprentissage
1) Apprentissages perceptifs
Ces apprentissages correspondent à la capacité de reconnaître et d’identifier des
stimuli déjà présents auparavant.
Ex :
● Reconnaissance d’info simples = niveau de brillance d’un signal lumineux.
● Reconnaissance d’info complexes = reconnaissance d’un visage
On pense que dans ce type d’apprentissage interviennent des structures et des
processus impliqués dans le traitement de l’info, qui sont différentes en fonction du
type d’info et aussi en fonction du niveau d’intégration de cas infos.
Simple = structures corticales qui traitent tôt l’info visuelle
Complexe = cortex visuel associatif.
Performance
Nombre
D’essais
Evolution rapide au début puis plateau

5
2) Apprentissages de type stimulus, réponse (I – R)
Le principe de base correspond à établir une relation entre un stimulus externe à
l’organisme et une réponse comportementale. Le sous jacent c’est l’établissement de
connexions nerveuses neuronales entre les structures de la perception et celles de la
programmation et/ ou de l’exécution motrice.
On connaît 2 types d’apprentissages :
L’apprentissage non-assiociatif :
Constitue la forme la plus simple d’apprentissage.
2 types :
l’habituation
Quand on répète une stimulation, la réponse à celle ci diminue progressivement en
amplitude jusqu'à une disparition totale ou partielle de réponse.
la sensibilisation
La réponse augmente avec la répétition des stimulations.
Ex : le réflexe de la pupille. Si on envoie un jet d’air il y a clignement.
Si le jet est faible elle clignera de moins en moins.
Si le jet est fort elle clignera de plus en plus.
L’apprentissage associatif :
C’est la mise en relation d’une stimulation à une réponse comportementale.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
1
/
25
100%