annale svt yahya

ANNALE TERMINALE S SCIENCE DE LA VIE ET DE LA TERRE
1 yahya MOHAMED MAHAMOUD

ANNALE TERMINALE S SCIENCE DE LA VIE ET DE LA TERRE
2 yahya MOHAMED MAHAMOUD
INTRODUCTION
Mise en contexte
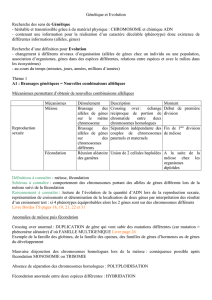
Approche du temps en biologie et géologie
Le déroulement des événements géologiques et biologiques sont caractérisés par des échelles
de temps, qui sont différentes en fonction des phénomènes étudiés.
L’histoire de la Terre, comme celle de la vie, s’inscrit dans celle de l’Univers. Elle fait appel à
de très longues durées. Une majorité d’astronomes estiment que l’Univers est âgé d’environ
15 milliards d’années. La Terre, elle, s’est formée il y a 4,6 milliards d’années et la vie y
est apparue il y a environ 3,8 milliard d’années. La planète et les êtres vivants ont alors
évolué conjointement.
Des événements majeurs jalonnent l’histoire de la Terre, dont la durée s’étale sur des
dizaines ou centaines de millions d’années, comme la mise en place de la tectonique des
plaques. Leur chronologie est établie à partir de la datation d’archives cosmiques,
géologiques ou biologiques.
Les êtres vivants ont laissé des traces au sein des roches. Une fois datés, ces fossiles
permettent de préciser l’histoire de la vie, qui est marquée par des étapes telles que
l’apparition de la cellule eucaryote ou du premier vertébré. Si la durée moyenne d’existence
d’une espèce est de l’ordre de quelques millions d’années, les durées de vie des êtres
vivants au sein des différentes espèces sont beaucoup plus courtes ; elles se mesurent en
années, en mois, en jours, voire en heures ou minutes (certaines bactéries peuvent se
diviser toutes les 30 minutes).La notion de temps à l’échelle des cellules est encore différente
; par exemple, un événement comme la mitose se mesure en heures.
Et à l’échelle des molécules, à l’intérieur même des cellules, ont lieu des réactions chimiques
extrêmement rapides : la plupart des enzymes peuvent catalyser de 103 à 106 réactions par
seconde.
_____________________________________
PARTIE I : PARENTE ENTRE ETRES VIVANTS
ACTUELLES ET FOSSILES-PHYLOGENETIQUES –
EVOLUTION.
_____________________________________
La recherche de parenté chez les vertébrés
En dépit de la formidable biodiversité attestée par les milliards d'espèces disparues et par les
quelques 2 millions d'espèces identifiées dans la nature actuelle, le monde vivant est avant
tout caractérisé par une remarquable unité. En effet, des caractéristiques fondamentales

ANNALE TERMINALE S SCIENCE DE LA VIE ET DE LA TERRE
3 yahya MOHAMED MAHAMOUD
partagées par tous les organismes révèlent une origine commune remontant à plus de 3,5
milliards d'années (Ga).
Tous les êtres vivants descendent d'un ancêtre commun et sont donc tous plus ou moins
apparentés, malgré les différences acquises au cours de l'évolution. C'est pour cette raison que
l'on s'efforce de les classer en fonction de leurs liens de parenté.
Ainsi, les divisions fondamentales de la classification au sein d'un règne (animal, végétal,
etc.) – les embranchements ou phylums – regroupent les espèces qui partagent un même plan
d'organisation. Il n'existe qu'un petit nombre de phylums (mollusques, arthropodes,
vertébrés…), mais on y trouve une multitude d'espèces différentes. Chacune d'entre elles est
caractérisée par des variations particulières du plan commun, résultant d'une évolution
différente des mêmes caractères ancestraux.
1. Comment déterminer les relations de parenté ?
Le principe utilisé consiste à identifier l'état ancestral d'un caractère et ses états dérivés
apparus au cours de l'évolution, de façon à remonter de proche en proche des descendants à
leur ancêtre commun.
Les caractères de même origine évolutive ou embryonnaires sont appelés caractères
homologues, même s'ils assurent parfois des fonctions différentes. Ils se distinguent des
caractères analogues qui, eux, assurent les mêmes fonctions sans avoir la même origine
évolutive, comme par exemple les ailes des insectes et celles des oiseaux.
Ainsi, tous les vertébrés tétrapodes (amphibiens, reptiles, oiseaux, mammifères) descendent
d'un ancêtre commun vieux de quelque 350 millions d'années. Ils possèdent en effet un même
plan d'organisation, caractérisé en particulier par un squelette interne comportant une colonne
vertébrale et deux paires de membres. Ces derniers sont des caractères homologues puisqu'ils
ont la même origine ; le membre antérieur, qui peut être une aile, un bras, une patte, une
palette natatoire, etc. a évolué différemment dans les quatre classes de vertébrés tétrapodes, à
partir d'une structure commune présente chez leur ancêtre commun.

ANNALE TERMINALE S SCIENCE DE LA VIE ET DE LA TERRE
4 yahya MOHAMED MAHAMOUD
2. Dans quels caractères peut-on reconnaître des homologies ?
On recherche des homologies aux différents niveaux d'organisation. Même si tous les êtres
vivants partagent une origine commune qui se traduit notamment par une remarquable unité
biochimique, génétique et cellulaire, les espèces se distinguent par des particularités
moléculaires, anatomiques, morphologiques, physiologiques, embryonnaires, larvaires,
comportementales, etc. La comparaison de ces différents types de caractères entre groupes
d'espèces appartenant au même phylum permet de reconnaître les homologies.
On a vu que les membres antérieurs des vertébrés tétrapodes sont homologues mais on peut
aussi identifier des homologies jusqu'au niveau moléculaire, notamment entre gènes ou
protéines. Ainsi, des protéines appartenant à des espèces différentes comme les globines
(hémoglobine, myoglobine) et les gènes qui les codent sont homologues.
Le degré d'homologie entre les caractères hérités d'un ancêtre commun est un marqueur du
degré de parenté entre les espèces. Pour les espèces fossiles, le nombre de caractères pris en
compte est forcément plus limité car, en général, seules les parties dures de l'organisme
(squelette, coquille) sont conservées. Dans certains cas favorables, on peut cependant aussi
étudier leur ADN (ADN fossile).
3. Comment évaluer le degré de parenté entre les espèces ?
Les séquences de molécules informatives — gènes et protéines — se prêtent particulièrement
bien à l'analyse informatique, ce qui permet de quantifier précisément leur degré de similitude
et donc de parenté.
Les gènes homologues dérivent d'un gène ancestral provenant de leur ancêtre commun. Plus
l'ancêtre commun est éloigné dans le temps, plus les gènes accumulent de mutations et plus ils
diffèrent par leur séquence. La comparaison deux à deux de ces séquences permet ainsi

ANNALE TERMINALE S SCIENCE DE LA VIE ET DE LA TERRE
5 yahya MOHAMED MAHAMOUD
d'évaluer la distance évolutive entre les espèces et de compléter les informations obtenues
par l'analyse d'autres caractères. Plus le nombre de caractères pris en compte est important et
plus les résultats sont précis. On peut ainsi reconstituer les filiations entre espèces.
4. Qu'est-ce qu'un arbre phylogénétique ?
Un arbre phylogénétique est un diagramme traduisant les relations de filiation entre les
espèces et leur plus proche ancêtre commun. Il faut cependant noter que ces ancêtres sont
hypothétiques puisque déduits de l'analyse des caractères homologues, par définition
différents du caractère ancestral.
On a pu ainsi construire l'arbre du vivant qui montre les relations phylogénétiques entre les
espèces et remonte de proche en proche jusqu'à l'origine de la vie, il y a plus de 3,5 milliards
d'années. La figure ci-dessous présente la partie de cet arbre correspondant aux reptiles, aux
oiseaux et aux mammifères, dont l'ancêtre commun hypothétique est daté de 350 millions
d'années.
5. Existe-t-il des fossiles d'ancêtres communs ?
Non, on ne peut pas trouver de fossile d'ancêtre commun car les notions de fossile et d'ancêtre
commun sont de natures différentes.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
1
/
47
100%