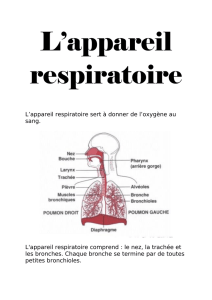
UV_102_BASES_PHYSIO_TRANSP_O2

1
BASES DE PHYSIOLOGIE
UV 102
TRANSPORT DE L’OXYGENE
ET DU CO2
P. PILARDEAU

2
I - TRANSPORT DE L’OXYGENE
1.1 GENERALITES
1.1.1 Composition de l’atmosphère
L’atmosphère est composée d’azote (79%), d’oxygène (21%) et de traces de gaz carbonique.
Chacun de ces gaz occupe une certaine fraction du volume total (exprimée en pourcentage) qui
correspond également à leur fraction de pression partielle.
La pression partielle de l’oxygène contenu dans l’air atmosphérique dépend de l’altitude. Au
niveau de la mer la PO2 est d’environ 150 mm Hg.
Variations de la PO2 en fonction de l’altitude et de la pression barométrique
Altitude Pression Pression
en mètres barométrique partielle en
en mm Hg oxygène (PO2)
0 760 160
1000 670 130
2000 596 115
3000 467 85
La pression partielle de chaque gaz est sensible à l’humidité de l’air. La vapeur d’eau occupe
en effet un certain volume et de ce fait représente une pression équivalente. Plus l’air est humide, plus
les pressions partielles en azote et en oxygène sont basses.
Au niveau des voies aériennes supérieures, l’air est saturé en vapeur d’eau. A 37°C cette
pression d’humidité (PH2O) correspond à 47 mm Hg.
1.1.2 Apport d’oxygène aux tissus
La fourniture d’oxygène aux tissus comprend quatre étapes :
= La ventilation pulmonaire
= La diffusion alvéolo-capillaire
= Le transport plasmatique
= La diffusion dans les tissus
1.2 - VENTILATION
La ventilation alvéolaire fonctionne suivant le système de la convection forcée. La différence
de pression entre l’air extérieur et l’air alvéolaire est due à l’action des muscles respiratoires. Au
repos seul le diaphragme est en activité. Sa contraction permet d’inspirer l’air (gradient de pression)
qui sera expulsé lors de la phase d’expiration de façon passive.

3
La fraction de gaz pulmonaire renouvelé à chaque inspiration au repos est relativement faible
(environ 10% de la totalité de l’air contenu dans les poumons. Ce mécanisme permet d’assurer une
grande stabilité de concentration de l’O2 et du CO2 pulmonaire et sanguin.
Ce mélange contient 14% d’O2 et 5,6% de CO2 correspondant respectivement à une pression
d’02 alvéolaire (PA O2) d’environ 100 mm Hg et de 40 mm Hg pour le CO2 (PA CO2)
1.3 - DIFFUSION ALVEOLO-CAPILLAIRE
Les échanges entre l’air alvéolaire et les gaz plasmatiques sont dus à la différence de
pression entre ces deux milieux.
Le gradient est très important pour l’O2 (60 mm Hg) et beaucoup plus faible pour le CO2 (5 à 6 mm
Hg).
Les échanges sont réalisés à travers une double couche cellulaire, celle de l’alvéole et celle du
capillaire, formée de :
= L’épithélium alvéolaire
= Une lame de tissu interstitiel
= L’endothélium vasculaire et sa lame basale.
Le total représentant environ 1. d’épaisseur.
CO2
Epithélium alvéolaire
Membrane basale
O2
Le gaz diffuse alors dans le plasma jusqu’au globule rouge. Seule une très faible fraction
d’oxygène reste dissout dans le sang (son rôle est cependant fondamental).
Au repos ce mécanisme est suffisamment actif pour n’utiliser que le premier tiers du capillaire
en contact avec l’alvéole.
Il existe de nombreux facteurs susceptibles de modifier la vitesse des échanges à ce niveau :
= Surface d’échange (surface de l’alvéole tapissée de capillaire). Plus elle est grande,
plus le transport est facilité.
= L’épaisseur de la paroi. Ce paramètre est relativement fixe chez le sujet non
pathologique.
= La vitesse de circulation du sang en rapport avec le débit cardiaque

4
PA O2 PA CO2
= 100 = 40
Pa O2 = 40 Pa CO2 = 46
La vitesse de transfert d’un gaz dépend de ses caractéristiques moléculaires. Pour un gradient
donné le transfert d’un gaz à travers la membrane dépendra de la capacité de ce gaz à traverser cette
membrane. Cette grandeur est différente d’un gaz à l’autre, elle est appelée capacité de diffusion.
Ainsi le CO2 présente une capacité de diffusion environ 20 fois plus forte que celle de l’oxygène.
La pression partielle mesurée dans le sang artérielle (80 à 100 mm Hg) est sensiblement plus
faible que celle des alvéoles (100 mm Hg). Ce phénomène est en rapport avec un shunt artério-
veineux physiologique (une fraction sanguine très faible ne passe pas par les capillaires
pulmonaires). C’est le cas pour un certain nombre de vaisseaux bronchiques et de veines
myocardiques qui se jettent directement dans les artères pulmonaires et le ventricule gauche.
Dans des conditions normales de pression atmosphérique, et en l’absence de phénomène
pathologique, l’hémoglobine est saturée entre 96 et 98% quand elle quitte les capillaires alvéolaire.
Les quelques pour cent manquant correspondent à l’effet des shuntes (hémoglobine court-circuitant le
passage pulmonaire). La monté en très haute altitude peut être désaturante, l’exercice physique n’est
pas désaturant.
1.4 - TRANSPORT PLASMATIQUE
L’O2 est transporté dans le sang sous deux formes :
= Dissoute
= fixée à l’hémoglobine
1.4.1 Oxygène dissout
La quantité d’un gaz dissout dans un liquide dépend de deux paramètres :
= La solubilité de ce gaz
= La pression partielle
L’oxygène étant un gaz très peu soluble, le contenu artériel en O2 dissout est donc très faible
(3%).
Si sur le plan quantitatif cette fraction joue un rôle négligeable dans le transport de l’oxygène,
elle joue à l’inverse sur le plan qualitatif un rôle majeur pour réguler les échanges
alvéole/hémoglobine et hémoglobine/tissus.

5
1.4.2 Hémoglobine
La majeure partie de l’oxygène est transportée par l’hémoglobine contenue dans les érythrocytes (ou
globules rouges).
1.4.2.1 La molécule d’hémoglobine
La molécule d’hémoglobine est formée de deux parties, l’hème qui fixera l’oxygène, et une
structure protéique appelée globine. Il s’agit d’une très grosse protéine qui, comme son nom l’indique
est « globulaire »,
1.4.2.1.1 L’hème
L’hème est formé de quatre noyaux pyrolles et d’un atome de fer ferreux lié aux quatre
atomes d’azote de la protoporphyrine. L’oxygène se fixe sur le fer.
CH3 CH =CH2
HC CH
H3C N CH3
N Fe N
HOOC - CH2 - CH2 CH = CH2
N
HC CH
HOOC - CH2 - CH2 CH
Un manque de fer (carence martiale) peut être à l’origine d’une anémie, et donc d’un trouble
du transport de l’oxygène (asthénie, essoufflement).
1.4.2.1.2 La globine
La globine est formée de quatre chaînes polypeptidiques identiques deux à deux , les chaînes
et .
Chaque chaîne est combinée à une molécule d’hème. Une molécule d’hémoglobine peut donc
fixer 4 oxygènes.
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%