psychologie et neuropsychologie cognitive i

Page 1 of 21
PSYCHOLOGIE ET NEUROPSYCHOLOGIE COGNITIVE I
Année académique 2010-2011
Titulaire: Philippe Mousty& Wim Gevers
PSYCHOLOGIE ET NEUROPSYCHOLOGIE COGNITIVE I ..................................................... 1
2. La cognition visuelle .................................................................................................................... 1
2.1 Mécanismes de saisie de l’information .................................................................................. 1
2.1.0. La lumière - qu'est-ce que c'est ...................................................................................... 1
2.1.1. Description anatomique de l’oeil humain ...................................................................... 2
2.1.2. Les voies visuelles de la rétine au cortex ....................................................................... 3
Types de cellules : ............................................................................................................ 4
Chiasma optique ............................................................................................................... 4
2.1.3. La rétine .......................................................................................................................... 4
2.1.4. Champs récepteurs des cellules ganglionnaires des CGL (corps Genouillées Latéraux
du Thalamus) ............................................................................................................................ 5
2.1.5. Patrons de réponses des cellules corticales (cellules du cortex : première zone de
projection de l’information) ..................................................................................................... 8
2.1.6. Architecture du cortex visuel primaire (V1) .................................................................. 8
2.1.7. Le cortex visuel extrastrié (V2-V5) ............................................................................... 9
2.1.8. Deux circuits anatomiquement et fonctionnellement distincts ? .................................... 9
2.2 La théorie de David Marr (1982) ......................................................................................... 10
2.3 Les processus perceptifs de bas niveau – Esquisse primale ................................................. 12
2.3.1 L’esquisse primale brute ............................................................................................... 12
2.3.2. L'esquisse primale complète......................................................................................... 12
2.3.3 L’esquisse 2 1/2-D (représentation intégrée des surfaces) ............................................ 14
2.3.3.1 La perception de la profondeur .............................................................................. 14
2.3.3.2 La perception du mouvement ................................................................................. 19
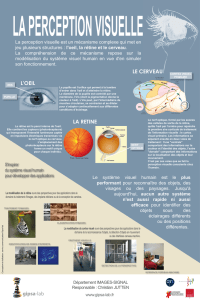
2. La cognition visuelle
On ne peut pas faire abstration des données biologiques : œil + circuit neuronaux.
2.1 Mécanismes de saisie de l’information
2.1.0. La lumière - qu'est-ce que c'est
Nous captons la lumière réfléchie par les surfaces
et les objets que l’on regarde.
La lumière est composée d’ondes
électromagnétiques (= flux de particules appelées
photons) dont la longueur varie. Onde sinusoïdale
longueur d’onde.

Page 2 of 21
Chaque longueur d’onde dans le spectre lumineux visible (380-780 nm) correspond à une
nuance de couleur allant du violet au rouge). On ne perçoit qu’une toute petite partie des
ondes.
Relations entre les caractéristiques physiques de la stimulation lumineuse et la perception:
Optique = ce qui est mesurable
Intensité (w) d’un signal lumineux
-> Luminance (cd/m²) (candela par m²)
Chrominance:
Longueur d’onde (nm)
Pureté (%)
Perception = ce que l’on voit
Luminosité (source lumineuse)
-> Brillance (objet brillant ou pas)
Chromaticité:
Teinte (nuance de couleur)
Saturation (vive : plus homogène, pâle)
2.1.1. Description anatomique de l’oeil humain
Cornée (protection extérieure)
Pupille : diaphragme (muscle) : permet de
régler la quantité de lumière qui pénètre dans
l’oeil
Cristallin (lens) : mise au point, accomodation
lentille (modification de la courbure, on
voit donc net plus ou moins loin)
Rétine : Tout le fond de l’œil, tapissé de nerfs
envoyés vers le nerf optique.
Nerf optique (blind spot = point aveugle,
disque optique)
Les yeux bougent à tout moment pour mettre
l'image la plus claire possible sur la tache jaune
(fovéa)

Page 3 of 21
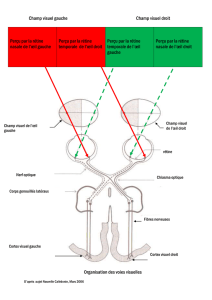
2.1.2. Les voies visuelles de la rétine au cortex
Au départ de l'oeil, les fibres nerveuses empruntent
soit la voie principale = voie géniculo-striée (vers les corps genouillés latéraux du
thalamus CGL/lateral geniculate nucleus; LGN) d'où elles se projettent sur le cortex
visuel primaire (ou strié, V1)
soit une autre voie sous-
corticale importante =
voie rétino-tectale vers
les colliculus supérieurs
(où elles sont en contact
avec des neurones
moteurs qui
commandent les
mouvements des yeux,
de la tête, du corps),
pulvinar -> cortex
visuel (pas V1) et autres
régions cérébrales:
Attention, mémoire,
contrôle du mouvement
occulaire.
Ces voies véhiculent de l'information dans les deux sens (feedback)

Page 4 of 21
Types de cellules :
photoréceptrices au niveau de la rétine (cônes et bâtonnets) :
détectent des points de lumière Transformation des signaux
lumineux en signaux nerveux.
ganglionnaires (Rétine et CGL): encodent la quantité de
lumière dans une région très délimitée de la rétine (plus
proche de la périphérie, elles ont des champs récepteurs
relativement larges càd qu'elles combinent l'information
provenant de larges aires de la rétine, plus proches de la
fovéa les champs récepteurs sont plus étroits. La
combinaison des réponses donne l'intensité moyenne de la
lumière)
corticales : certaines détectent les lignes et les contours, d'autres contribuent à des processus
plus élaborés (reconnaissance des formes et objets) différents rôles
NB: il y en a des binoculaires, que le stimulus soit présenté à un oeil ou à l'autre, elles
réagissent.
Chiasma optique
Croisement des fibres nerveuses issues des faces rétiniennes
nasales de l'oeil
NB: chaque rétine est divisée en deux hémirétines: une nasale
interne et une temporale externe, les voies optiques des
hémirétines nasales subissent un chgmt de côté au niveau du
chiasme
ipsilatéral: du même côté du corps, contralatéral : du côté opposé
Chaque hémichamp visuel comprend le champ nasal de l'oeil
contralatéral et le champ temporal de l'oeil ipsilatéral. Les
champs visuels temporaux étant plus étendus que les champs
visuels nasaux, la vision de la périphérie du champ visuel est
purement monoculaire.
HVG est vu par nasal G qui est croisé à D et temporal D qui reste
à D => tt va ds HD
=> représentation contralatérale de l’espace visuel au niveau du cortex : Hémichamp visuel
gauche (HVG) -> hémisphère Droit (HD), et inversément: HVD-> Rétine G -> HG
2.1.3. La rétine
plus de 10 sortes de cellules différentes, 3 couches
+- 127 millions de cellules
photoréceptrices (par oeil) dans le
fond de la rétine

Page 5 of 21
+- 800.000 cellules ganglionnaires (par oeil) envoient les infos vers le cerveau
Les cellules ganglionnaires concentrent donc l’activité de nombreux photorécepteurs
Ce phénomène de convergence est nettement moins marqué pour
les cônes de la fovéa qu'en périphérie
Une cellule ganglionnaire est un type de neurone situé dans la
rétine de l'œil qui reçoit une information visuelle des
photorécepteurs via de nombreux intermédiaires cellulaires tels
que les cellules bipolaires, les cellules amacrines, et les cellules
horizontales. Les axones des cellules ganglionnaires de la rétine
sont myélinisés. La partie myélinisée se situe en dehors de l'œil.
Ces axones forment le nerf optique et sont connectés au corps
genouillé latéral dans le cerveau. Les cellules ganglionnaires de la
rétine envoient des axones au corps géniculé latéral via le nerf
optique.
Deux types de cellules photoréceptrices :
1. Cônes ( 7 millions):
Vision des couleurs (3 types de cônes: rouge, vert et bleu)
Faible sensibilité (-> vision diurne, photopique = de jour)
Haute résolution spatiale, grande acuité
Principalement concentrés dans les régions fovéale (rouge et vert) et parafovéale =>
perception des détails fins
2. Bâtonnets (120 millions):
Vision achromatique (pigment visuel = rhodopsine) Pas sensible à la longueur d’onde.
Faible résolution spatiale, grande sensibilité (vision scototopique - de nuit), très grande
résolution temporelle
Distribués sur toute la surface de la rétine (sauf fovéa)
perception ambiante, globale, détection des mouvements
La lumière engendre des réactions chimiques au niveau des pigments visuels qui sont traduites
par la cellule en activité électrique
œil gauche: Distribution des cônes
(ligne) et bâtonnets (pointillés) sur la
rétine par rapport à la fovéa, nombre
de cellules par mm² (bleu = tache
aveugle)
2.1.4. Champs récepteurs des cellules
ganglionnaires des CGL (corps Genouillées Latéraux du Thalamus)
Kuffler (1953) enregistre l’activité de cellules ganglionnaires chez le chat, ces cellules ne
réagissent que si une région bien précise du champ visuel est stimulée (champ récepteur). Il met
donc en évidence deux types de cellules dont les champs récepteurs concentriques diffèrent :
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%