I. Les parentés permettent de reconstituer une histoire évolutive

RR - 15/04/17 - 769771286 - 1/7
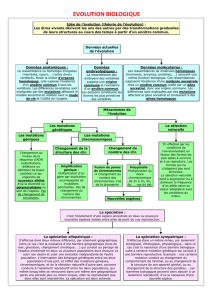
Parentés et évolution
I. Les parentés permettent de reconstituer une histoire évolutive
A. Les caractères homologues sont hérités d'un ancêtre commun
B. La phylogénie reconstitue une histoire évolutive
►TP 1. Parentés anatomiques
II. Les caractères homologues sont macroscopiques ou moléculaires
A. À l'échelle macroscopique on distingue l'état ancestral et l'état dérivé d’un caractère
B. À l'échelle moléculaire le degré de similitude permet de préciser parentés
III. L’évolution repose sur des mécanismes génétiques
A. Des accidents génétiques sont source d’innovation
►TP 2a. Polyallélisme et mutations (version Anagène)
►TP 2b. Polyallélisme et mutations (version papier)
B. Les innovations génétiques ont des succès divers
► TP 3. Génotype et environnement
OBJECTIF
► FIGURE 1. Unité du vivant rappels 2de et 1ère ES d’après Belin TS p. 18
Les êtres vivants partagent des propriétés communes (structure
cellulaire, universalité de la molécule d'ADN, du code génétique et de
l’expression des gènes, plan d'organisation génétiquement déterminé,
etc.) qui traduisent des liens de parenté.
Les organismes actuels et fossiles résultent de l'évolution, c'est à dire de
la transformation d'espèces au cours du temps.
On cherche à préciser les relations de parenté entre les vertébrés
ainsi que les principaux mécanismes de l’évolution.
N.B. Selon la terminologie actuelle, le nom des groupes et les noms vernaculaires des
espèces s’écrivent en minuscules.
Première ES
Chapitre
7.1
4 semaines

RR - 15/04/17 - 769771286 - 2/7
I. Les parentés permettent de reconstituer une histoire
évolutive
A. Les caractères homologues sont hérités d'un ancêtre commun
On appelle caractères homologues des structures (des attributs) qui présentent
la même organisation générale dans plusieurs espèces, cela relève d'une
similitude de contrôle génétique.
► FIGURE 2. Le membre antérieur des vertébrés dans Bordas TS p. 20.
► VOIR aussi Bordas ES p. 288 (figure équivalente).
Exemple. Les membres antérieurs des vertébrés sont homologues, car ils
présentent la même organisation (bras (humérus), avant bas (radius + cubitus),
carpe (poignet), métacarpe (paume), phalanges (doigts)).
Les ailes d’oiseau et de chauve-souris sont homologues en tant que membres mais
ne sont pas homologues en tant qu’ailes car, de ce point de vue, leur organisation est
différente.
L'aile d'oiseau et celle d'insecte ne sont pas homologues car elles n’ont pas la même
organisation (simple convergence de forme, il y a analogie).
Des structures homologues (membre antérieur des vertébrés) peuvent avoir une forme
ou une fonction différente (marche, nage ou vol), alors que deux structures analogues
(ailes d’oiseau et d’insecte) peuvent avoir la même fonction (vol).
►FIGURE 3. Un exemple de relations de parenté RR, voir aussi Nathan TS p.
33.
Un caractère homologue possédé par un groupe d'espèces (les vertèbres pour les
vertébrés), est hérité d'une population ancestrale commune ou ancêtre
commun chez qui l'innovation évolutive est apparue.
B. La phylogénie reconstitue une histoire évolutive
Un arbre phylogénétique indique les caractéristiques du plus récent ancêtre
commun d'un ensemble d'organismes :
- chaque nœud correspond à un ancêtre commun hypothétique ;
- chaque extrémité de branche porte un organisme actuel ou fossile ;
Un arbre phylogénétique est construit à partir d'une matrice de caractères (exprimée
en présence ou absence d’une série de caractères dans une série d’espèces
Les ancêtres communs ne correspondent pas à des espèces fossiles ou actuelles
précises, qui ne sont jamais placées aux nœuds d'un arbre phylogénétique. En effet on
ne connaît pas la descendance des espèces fossiles ou actuelles. Un arbre
phylogénétique (qui est plus proche de qui) n’est donc en rien un arbre généalogique
(qui descend de qui).
►FIGURE 4. Arbre phylogénétique des vertébrés dans Bordas p. 27. Voir aussi
Nathan p. 35 et fiche agrég RR d'après La Recherche n°117 décembre 1980.
ATTENTION. Ne pas tenir compte de la figure du Bordas ES p. 292.
Les organismes qui dérivent d'un même nœud partagent des caractères
communs exclusifs, il sont apparentés et forment un groupe monophylétique
(= clade).
Exemples : eucaryotes, vertébrés, mammifères, primates…
Exemple. Les vertébrés forment un groupe monophylétique car ce groupe
réunit exclusivement tous les descendants d'un ancêtre commun chez qui
apparaît une nouveauté évolutive : les vertèbres.
Un groupe monophylétique se définit, sur le plan phylogénétique, par ce que ses
membres ont en commun (ex. des vertèbres pour les vertébrés) et non par ce qu’ils ne

RR - 15/04/17 - 769771286 - 3/7
possèdent pas (ex. pas de vertèbres, les invertébrés ne forment pas un groupe
phylogénétique). En effet l’absence de caractère est infinie, quelle que soit l’espèce
considérée.
L'ancêtre commun à tous les vertébrés est donc plus proche (= plus récent) que
l'ancêtre commun aux vertébrés et aux insectes. Les vertébrés sont plus apparentés entre
eux qu'avec les insectes.
On appelle groupes frères, deux groupes qui descendent d'un même ancêtre
commun immédiat et possédant les mêmes caractères dérivés exclusifs (myxines
et vertébrés, crocodiles et oiseaux, homme et chimpanzé).
Un arbre phylogénétique permet de situer chronologiquement les une par
rapport aux autres les diverses innovations évolutives (= chronologie relative
c’est à dire sans dater).
►TP 1. Parentés anatomiques
► VOIR. Correction du TP 1
II. Les caractères homologues sont macroscopiques ou
moléculaires
Les caractères homologues s’observent à différentes échelles :
- macroscopique : accessible à l’œil nu (caractères embryonnaires, morphologiques (=
organisation externe), anatomiques (= organisation interne)) ;
- microscopique (= cellulaire) accessible au microscope et qui est abordée ici comme
l’échelle macroscopique ;
- moléculaire accessible à partir d’une analyse chimique.
A. À l'échelle macroscopique on distingue l'état ancestral et l'état
dérivé d’un caractère
Dans un ensemble d’espèces, un même caractère peut se trouver sous plusieurs
états. Dans une espèce il peut être non modifié par rapport à un ancêtre lointain,
on parle d'état ancestral, ou modifié dans d’autres, on parle d'état dérivé.
MATERIEL. Squelette humain, main de cheval (ou de vache) et crâne de cheval
(ou de vache).
Exemple. L'homme possède 5 doigts à la main et au pied, comme les premiers
vertébrés sortis des eaux il y a 350 Ma. C'est un état ancestral. Le cheval
possède un seul doigt, c'est un état dérivé. Inversement, l'homme présente une
régression de la face, qui est un état dérivé alors que le cheval possède un
museau, qui est un état ancestral. Chez l'homme et le cheval la main et la face
n'ont pas évolué à la même vitesse.
L'état ancestral d'un caractère est toujours recherché dans un extra-groupe, c'est à
dire un groupe extérieur à l'échantillon utilisé. L’état ancestral a été acquis avant
l’ancêtre commun, alors que l’état dérivé n’a été acquis qu’à partir de l’ancêtre
commun.
Le partage des mêmes états dérivés pour plusieurs caractères témoigne d'un
ancêtre commun proche, c'est donc le signe d'une parenté étroite. Inversement le
partage d'un caractère ancestral ne témoigne que d'une parenté éloignée.
Exemple. L'homme est plus apparenté avec le cheval qu'avec la tortue car il
possède avec le cheval une série de caractères dérivés mammaliens, et avec la
tortue un caractère ancestral (5 doigts à la main et au pied). L'ancêtre commun à
l'homme et au cheval est donc plus récent que l'ancêtre commun à l'homme, au
cheval et à la tortue.

RR - 15/04/17 - 769771286 - 4/7
Quand deux lignées divergent à partir d’un ancêtre commun, elles continuent, chacune
de son côté, à évoluer de manière indépendante. L’ancêtre commun à l’homme et au
cheval n’est donc ni un homme, ni un cheval et ce dernier n’est pas l’ancêtre de
l’homme. De la même manière l'ancêtre commun à l'homme et au gorille nous est plus
proche que l'ancêtre commun à l'homme et au cheval car l'homme partage des caractères
dérivés avec le gorille (réduction de la face, pouce opposable, ongles plats...). Cet
ancêtre commun n’était ni un homme, ni un gorille.
Le principe de parcimonie (= principe d’économie d’hypothèses), qui est utilisé pour
bâtir les arbres phylogénétiques, postule qu'une même nouveauté évolutive (exemple la
réduction de la face) n'apparaît qu'une seule foi. Cela reste souvent à démontrer.
B. À l'échelle moléculaire le degré de similitude permet de préciser
parentés
Seules les molécules séquencées (acides nucléiques et protéines) peuvent être
utilisées pour déterminer des phylogénies. Les autres molécules (glucidiques et
lipidiques) sont communes à beaucoup d’espèces (glucose, glycogène, amidon,
cholestérol…) et sont inutilisables ici.
►FIGURE 5 . La rhodopsine dans le règne animal RR d’après Phylogène, voir
pour précisions Nathan TS p.25. Les chéloniens sont les tortues.
La forte ressemblance de séquence de deux molécules appartenant à deux ou
plusieurs espèces souligne leur homologie. Elles dérivent d'une molécule
possédée par le plus récent ancêtre commun aux espèces envisagées.
À partir de données moléculaires, un arbre phylogénétique repose sur une matrice
des distances qui est un tableau à double entrée exprimant, pour une série d’espèces
prises deux à deux, le nombre de différences (ou de ressemblances) dans des séquences
homologues de nucléotides ou d’acides aminés.
► FIGURE 6. Comparaisons moléculaires RR d’après Phylogène.
Au niveau des mutations il y a un risque élevé de réversion (une mutation annule
une mutation précédente) et de convergence. En fait, à l'échelle moléculaire, le
caractère est le nucléotide ou l'acide aminé et non la molécule. On n'utilise donc pas
les termes "état primitif" et "état dérivé" pour les données moléculaires.
III. L’évolution repose sur des mécanismes génétiques
Gène : partie de molécule d'ADN qui constitue l'unité d'information gouvernant la
synthèse d'une protéine donnée.
Allèle : une des versions d’un gène.
Génome : ensemble de l'information génétique portée par les chromosomes d’un
individu ou d’une espèce. Les génomes actuels gardent en archive des innovations
génétiques survenues au cours du temps.
A. Des accidents génétiques sont source d’innovation
1. Les mutations permettent l’apparition de nouveaux allèles
Une mutation est une modification au hasard de la séquence des nucléotides
d'un gène (substitution (= remplacement), délétion (= perte), insertion (= ajout)).
Elle est typiquement spontanée (= aléatoire), et non orientée (= sa nature ne
dépend pas des caractéristiques du milieu).
Les mutations sont plus nombreuses sous l'action de certains agents mutagènes comme
les rayons ionisants (rayons X et gamma), les ultraviolets (voir cours de 2de chapitre

RR - 15/04/17 - 769771286 - 5/7
2.2.TP3 et §II.C) ou certaines substances chimiques (benzène...). Ces agents augmentent
la fréquence des mutations mais n'orientent pas leur nature.
Généralement, une mutation permet seulement l'apparition d'une nouvelle version
d'un gène, ou allèle. Elle n'entraîne donc pas l'apparition d'une nouvelle espèce,
mais introduit seulement de la diversité allélique au sein de l'espèce.
Une mutation peut intervenir lors de la réplication de l'ADN qui précède toute
division cellulaire (mitose ou méiose) ou résulter d'un remaniement chromosomique.
La fréquence des mutations (= taux de mutation) est faible pour un gène donné.
Elle est de l'ordre de 10-6 (variable selon les organismes et les gènes de 10-3 à 10-11).
Cependant, compte tenu du grand nombre de gènes (environ 35 000 chez l'homme) la
probabilité d'une mutation par gamète n'est pas négligeable (environ 1%).
Pour un gène donné, la rareté des mutations permet le maintien des caractéristiques de
l'espèce pendant quelques centaines de générations. Cela n'empêche pas le polyallélisme
et donc une variabilité intraspécifique pour de nombreux caractères.
►TP 2a. Polyallélisme et mutations (version Anagène)
►TP 2b. Polyallélisme et mutations (version papier)
Selon le temps disponible et la motivation des élèves le TP2 se fait sous forme
de TP(TP2a), de TD (TP2b) ou, plus simplement en ES, en support du cours
(transparents de TP2b).
2. Les duplications permettent la multiplication des gènes
► FIGURE 7. Un mécanisme d’apparition d’un nouveau gène RR
On appelle duplication génique, une mutation particulière qui provoque
l’apparition d’une copie supplémentaire d’un gène entier.
Les mécanismes de duplication génique sont hors programme.
Ces gènes copiés peuvent rester groupés sur le même chromosome ou bien se
disperser sur deux ou plusieurs chromosomes différents (= transposition).
3. Des mutations sur des gènes dupliqués entraînent l’apparition de
nouveaux gènes
► FIGURE 8. La famille multigénique des globines (au complet) dans Ridley,
De Boeck, 1997 voir aussi Nathan TS p. 78.
Les gènes dupliqués peuvent :
- ne pas muter (nous possédons ainsi des milliers de gènes identiques d'histone
(protéines associées à la structure des chromosomes) ou d'ARN ribosomal), on
parle de gènes répétés ;
- muter chacun pour son compte, tout en restant apparentés, les nouveaux
polypeptides codés conservent la même fonction (cas des globines). On appelle
famille multigénique un ensemble de gènes issus d'un même gène ancestral.
Dans ce cas une mutation délétère est sans conséquence car une copie intacte du gène
initial est conservée.
- muter chacun pour son compte en ne conservant pas la même fonction. Les
nouveaux gènes permettent alors l'acquisition de nouvelles fonctions (cas des
hormones post-hypophysaires ou de la myoglobine (stockage de O2) par rapport
aux autres globines (transport de O2)).
Certains gènes dupliqués, les pseudogènes, ne sont plus fonctionnels.
On appelle gènes chimères des gènes formés par la réunion de deux ou plusieurs
séquences d’ADN qui seront transcrites en une seule unité.
On estime qu'une similitude (même acide aminé situé à la même position) entre
deux protéines supérieure à 20 % indique une parenté, c'est à dire une origine commune.
 6
7
6
7
1
/
7
100%