Comment la glycémie est elle régulée

Comment la glycémie est elle régulée ?
La glycémie, quantité de glucose dans le sang, est un facteur primordial pour un bon
état de santé. Bien que les apports de glucose soient très variables dans le temps, la
glycémie reste toujours comprise, chez l’être humain sain à jeun, entre 4,5 et 6,5 mmol/l. Cette
régulation est assurée par les sécrétions endocrines du pancréas qui pénètrent dans le flux
sanguin par la veine mésentérique.
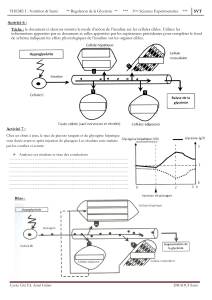
Les principales hormones d'origine pancréatique sont au nombre de trois:
- l'insuline: synthétisée par les cellules B, qui est une hormone hypoglycémiante.
- le glucagon: synthétisée par les cellules A, est une hormone hyperglycémiante. C'est
un facteur antagoniste de l'insuline. Il agit en stimulant la glycogénolyse hépatique. On
parle alors d’hormone du jeûne.
- la somatostatine: synthétisée par les cellules D, qui est aussi une hormone
hyperglycémiante.
Ces hormones n’agissent pas seulement sur le métabolisme glucidique mais exercent
également une profonde influence sur le métabolisme des lipides et des protides. Une
perturbation majeure de leur production ou de leur équilibre dynamique, telle que le DNID, va
donc avoir des retentissements très généraux et concerner l’ensemble du métabolisme.
On pourrait dire que la régulation de la glycémie commence déjà dès que de la
nourriture est vue, caractérisant un réflexe conditionné! En effet, des influx nerveux d’origine
cervicale mais également œsophagienne (associés à la déglutition) stimulent, la libération de
l’insuline par les cellules B grâce aux terminaisons issues du nerf pneumogastrique droit
libérant de l’acétylcholine. Le pancréas est ainsi prévenu par avance de l’arrivée imminente de
nourriture dans le duodénum. Au niveau stomacal, une molécule cosécrétée avec l’insuline,
l’IAPP, ralentit le passage de la nourriture entre l’estomac et l’intestin. De ce fait, si l’organisme
est nourri “en continu” avec pour conséquence une insulinémie assez élevée, les aliments
subissent une rétention gastrique qui empêche l’arrivée d’une trop grande quantité de glucose
dans l’intestin.
Dès que le contenu stomacal pénètrent dans le duodénum, celui-ci envoie de
nombreux signaux hormonaux qui prennent leur origine dans les cellules endocrines de la
paroi intestinale. Certaines hormones de l’intestin, comme la somatostatine, inhibent la

sécrétion stomacale, d’autres, telles que la sécrétine, l’entéroglucagon ou la gastrine, stimulent
les sécrétions endocrines pancréatiques. Le GIP (Gastric Inhibitory Peptide) cumule ces deux
effets.
Le glucose du chyle est activement absorbé en même temps que les ions sodium au
niveau des microvillosités des entérocytes. Une perméase permet ensuite le rejet du glucose
absorbé au pôle basal de ces cellules, accompagnée d’un rejet actif d’ion sodium et d’une
entrée de potassium dans la cellule.
Ce sont les oses comme le glucose, mais aussi le fructose, le ribose, le galactose et le
mannose qui stimulent fortement la sécrétion et la libération d’insuline par les cellules B (Le foie
transformant in vivo fructose et galactose en glucose). Les acides aminés issus de la digestion
des protéines stimulent eux aussi la production de cette hormone. Simultanément, l’insuline
produite inhibe localement (effet paracrine) la production du glucagon. On retrouve donc dans
la veine porte une forte concentration en glucose et en insuline.

Chronologiquement, le premier effet de l’insuline va être de favoriser au niveau
hépatique la transformation du glucose sanguin en glycogène mis en réserve dans les
hépatocytes. Après le foie, l’insuline va agir sur les cellules musculaires qui vont capter le
glucose sanguin et le transformer, elles aussi, en glycogène. Les adipocytes, à leur tour,
verront leur capture du glucose ainsi que sa transformation en triglycérides stimulée par
l’insuline.
La pénétration du glucose dans les cellules est liée à la présence à la surface de
celles-ci de récepteurs cytoplasmiques.
Il existe différents types de récepteurs; et l’insuline agit en provoquant l’exocytose de
récepteurs GLUT 4 au niveau des cellules qui en sont dotées: on est donc conduit à différencier
des tissus non insulino-dépendants (cerveau, rétine, érythrocytes) et des tissus insulino-
dépendants (le foie, le muscle et le tissu adipeux) dont les cellules possèdent le GLUT 4.

L'expression de ces récepteurs à la surface des cellules est modulée par le taux sanguin
d'insuline. L’entrée du glucose dans les cellules de tissus insulino-dépendants provoque une
diminution progressive de la glycémie après le repas. Cette diminution entraîne par rétro-
action une baisse de la sécrétion d’insuline, qui est détruite en continu au niveau hépatique par
une enzyme, la glutathion insuline transhydrogénase, qui rompt les ponts disulfures reliant les
deux chaînes A et B de cette molécule.
Dans l’organisme, le glucose est utilisé par toutes les cellules: il leur fournit une source
d’énergie importante libérée dans une chaîne de réactions de dégradation faisant intervenir de
l’ATP jusqu’a obtention d’acide lactique. Cependant, la plupart des tissus peuvent utiliser
également à cette fin les corps cétoniques. Le glucose constitue aussi une base de synthèse
des acides gras et triglycérides. Le premier stade de métabolisation du glucose dans la cellule
est la formation du glucose 6 phosphate (G6P) grâce à l’enzyme hexokinase. Cette molécule
sert de substrat à 4 autres enzymes:
- une isomérase qui catalyse la transformation du G6P en G1P pour donner finalement
du glycogène
- une enzyme permettant la formation de fructose 6 phosphate, engageant ainsi une
biodégradation par glycolyse anaérobie aboutissant à la formation d’acide pyruvique.
- une G6P déshydrogénase permettant la transformation du G6P en
phosphogluconolactone dégradé par la voie des pentoses.
- une glucose 6 phosphatase hépatique permettant la libération du glucose et son
passage dans le sang en cas de besoin.
Suite à l’action de l’insuline, la glycémie peut descendre en deçà de la normale: c’est
alors le glucagon, synthétisé par les cellules A, qui stimulera au niveau hépatique la
glycogénolyse et surtout la néoglucogénèse apte à résorber pendant plusieurs heures toute
tendance hypoglycémique. Pour cela, le glucagon se fixe a un récepteur membranaire
hépatique, et via une protéine kinase AMPc dépendante, il stimule la glycogène
phosphorylase et inhibe parallèlement la glycogène synthase.
Chez les mammifères, l'organisme contrôle donc l'équilibre entre la consommation
cellulaire du glucose, sa production endogène hépatique et les apports exogènes. Les
facteurs hormonaux comme l'insuline et le glucagon, des neurotransmetteurs ainsi que d'autres
molécules participent à la régulation du métabolisme glucidique afin de maintenir l'homéostasie
(fig. C). Certains des signaux hormonaux favorisant la production d'insuline ont été étudié
dans le but de corriger le DNID. C'est le cas en particulier du GIP (Peptide Inhibiteur
Gastrique) qui a donné des résultats décevants, alors que le "glucagon intestinal" (GLP-1)
semble plus prometteur.
Les régulations des sécrétions endocrines pancréatiques font donc intervenir de
nombreuses molécules d'origines diverses: des hormones, mais aussi des
neurotransmetteurs et des produits de la digestion.

1
/
5
100%