Du petit pois à la génétique humaine - Site SVT - Aix

8
NEPHROGÈNE N°34
Avril 2003
Mendel 1,
le père naturel de
la génétique
Quand, en 1900, De Vries, Correns
et von Tschermak constatent que
leurs expériences de croisements
chez le pois, le maïs, le chrysanthè-
me, la véronique et bien d’autres
plantes donnent des résultats iden-
tiques à ceux qu’un moine morave,
Gregor Mendel, avait publiés en
1865, ces trois botanistes réalisent
qu’une théorie de l’hérédité était née
depuis 35 ans sans que personne, ni
eux mêmes, ni même Mendel, n’en
ait été conscient. De cette “redécou-
verte” des lois de Mendel est née la
légende du “génie incompris et
oublié”. Légende, car il n’y a rien de
bien mystérieux à cela. En effet,
Mendel n’a jamais considéré qu’il
recherchait les lois générales de l’hé-
rédité. Il était simplement concerné
par deux questions, l’une théorique,
l’autre pratique.
●
la question théorique était de
savoir si chacun des sexes jouait
un rôle dans la procréation (pour
Aristote, tout est déterminé par le
sexe mâle, la femelle ne jouant
qu’un rôle de matrice) ; pour cela,
Mendel va démontrer par l’expéri
mentation que les contenus des
cellules sexuelles ou gamètes, aussi
bien mâles que femelles, jouent un
rôle dans l’hérédité.
●
la question pratique est celle de la
stabilité des croisements ou
“hybrides”. Depuis deux siècles
, les
“hybrideurs” souhaitaient pouvoir
croiser des souches pures afin
d’obtenir des hybrides stables,
ayant par conséquent un intérêt
agronomique. Par ses expériences
sur le pois, Mendel va montrer que
les croisements peuvent donner
une multitude de descendants
hybrides parmi lesquels certains,
assez rares, question de probabi-
lité, peuvent être stables ; il crée
lui-même plusieurs hybrides
stables de fuschia et apporte ainsi
une méthode pratique aux hybri-
deurs qui souhaitent construire de
nouvelles variétés par croise-
ments.
Mais à aucun moment Mendel n’a
pensé que les lois de l’hérédité chez
le pois pouvaient avoir une portée
universelle dans tout le règne vivant,
incluant végétaux, champignons et
animaux, dont l’homme évidem-
ment. Mendel a conçu la génétique,
sans le savoir, il en est le père
naturel.
Ce qu’a découvert
Mendel et ce qui s’en
suivit !
Mendel a une vision des phéno-
mènes naturels venant de sa
formation de physicien et appuyée
sur des principes rationalistes. Elle
consiste d’abord à concevoir deux
univers, celui des effets et celui des
causes qui les déterminent, puis à
considérer que l’on peut entre-
prendre l’observation d’un phéno-
mène assez simple pour qu’il soit
l’effet d’une cause unique.
C’est en se fondant sur cette double
démarche qu’il va réinterpréter les
résultats des croisements de varié-
tés de pois (Pisum sativum) connus
de tous les hybrideurs botanistes
depuis les années 1820.
Suivons Mendel dans son raisonne-
ment, en en profitant pour définir les
termes de la génétique moderne à la
naissance de laquelle le destin ne lui
a pas permis d’assister.
1. Je dispose de variétés pures de
pois, l’une à fleurs mauves, l’autre
à fleurs blanches. Ces variétés
sont pures, car lorsqu’on les laisse
se reproduire naturellement (par
autofécondation), elles donnent
toujours des descendants iden-
tiques aux parents : à fleurs
mauves chez la première, à fleurs
blanches chez la seconde.
DU PETIT POIS DE MENDEL
A LA GÉNÉTIQUE HUMAINE
DU PETIT POIS DE MENDEL
A LA GÉNÉTIQUE HUMAINE
■
Professeur Jean-Louis Serre,
Laboratoire de cytogénétique
et génétique moléculaire humaine,
Université de Versailles.
Saint Quentin en Yvelines,
Versailles.

9
NEPHROGÈNE N°34
Avril 2003
Je m’intéresse à une caractéristique
observable du pois, sa couleur, que
mes successeurs, les généticiens,
définiront comme un caractère.
Ce caractère se présente sous deux
formes qu’on appellera phénotype,
le phénotype mauve et le phénotype
blanc. Mes successeurs diront qu’un
caractère se présente sous plu-
sieurs formes appelées phénotypes.
2. Si je considère que la différence
de couleurs entre les deux
variétés ne dépend que d’une
seule cause, je déclare que cette
cause, ou ce facteur, existe sous
deux formes : la forme A, respon-
sable de la couleur mauve et la
forme a, responsable de la couleur
blanche. Mes successeurs donne-
ront à ce facteur le nom de gène
et les deux formes de ce gène,
impliquées dans la réalisation des
deux phénotypes, seront appe-
lées allèles du gène.
3. Je réalise des croisements entre
ces deux variétés en procédant à
la castration des anthères (ces
organes formant le pollen sont les
gamètes mâles) afin de féconder
les stigmates (ces organes renfer-
ment les gamètes femelles) avec
du pollen venant de l’autre varié-
té. Je réalise de la sorte deux
croisements dits réciproques :
dans un cas, le pollen est issu
d’une plante à fleurs mauves alors
que dans l’autre il est issu d’une
plante à fleurs blanches.
4. J’observe que toutes les plantes
issues de ces deux croisements
ont des fleurs mauves et qu’auc
ne n’a de fleurs blanches.
a. je peux déjà conclure que, si
seul un des deux sexes impor-
tait, le mâle ou le femelle, alors
les descendants d’un croise-
ment devraient être blancs et
ceux de l’autre croisement
devraient être mauves, ce qui
n’est pas le cas, donc Aristote
avait tort…
b. je dois aussi conclure que
toutes les fleurs étant mauves,
le phénotype mauve est dit
“dominant” et que le phéno-
type blanc est dit “récessif”.
c. mais je n’ o b s e rve que les
phénotypes, c’est-à-dire l’effet
des causes, et je sais que ces
causes sont aussi bien trans-
mises par le pollen que par
l’ovule. La question est de
savoir si, malgré leur phénotype
mauve, les individus issus des
croisements ne renferment que
des allèles A responsables du
phénotype mauve, ou si ils peu-
vent aussi renfermer des allèles
a dont l’effet causal ne pourrait
pas se manifester concurrem
ment à celui de A, ce qui
expliquerait alors pourquoi seul
le phénotype mauve est
présent à cette génération.
5. Pour le savoir, je laisse ces plantes
de première génération fabriquer
leurs propres gamètes, puis
s’auto-féconder et je plante les
pois issus de cette autoféconda-
tion et je constate que 75 %
des plantes de deuxième généra-
tion ont des fleurs mauves alors
que 25 % ont des fleurs
blanches.
a. je peux donc conclure que
l’allèle a était présent dans les
plantes mauves de première
génération puisqu’on retrouve
son effet, le phénotype blanc,
dans les plantes de seconde
g é n é r a t i o n . L e s p l a n t e s
mauves de première génération
renferment à la fois A et a :
m e s s u c c e s s e u r s d i r o n t
qu’elles sont h é t é r o z y g o t e s ,
car elles contiennent deux
allèles différents du gène
concerné par l’analyse.
b. je peux même faire l’hypothèse
que chaque gamète fabriqué
par l’hétérozygote Aa est “pur”
et ne renferme qu’un seul des
deux allèles, A avec une proba-
bilité égale à 50 %, ou bien de a
avec une probabilité égale à
50 %, de sorte que l’union au
hasard des gamètes, lors de
l’autofécondation, conduira à
25 % de AA, 50 % de Aa et
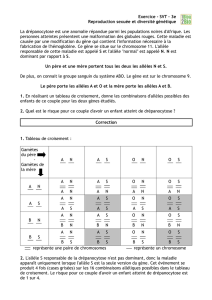
25 % de aa, selon le tableau de
croisement ci-dessous, soit
exactement ce qui est observé,
75 % de plantes à fleurs
mauves et 25 % de plantes à
fleurs blanches.
Pollen A a
Ovule
A AA Aa
mauve mauve
a Aa aa
mauve blanc
À LA GÉNÉTIQUE HUMAINE
À LA GÉNÉTIQUE HUMAINE
DU PETIT POIS DE MENDEL
DU PETIT POIS DE MENDEL

10
NEPHROGÈNE N°34
Avril 2003
À LA GÉNÉTIQUE HUMAINE
À LA GÉNÉTIQUE HUMAINE
DU PETIT POIS DE MENDEL
DU PETIT POIS DE MENDEL
c. je peux même, si ma théorie
est juste, prédire que mes
plantes à fleurs blanches sont
pures car elles contiennent
deux allèles identiques du
gène, ce que mes successeurs
désigneront comme homozy-
g o t e s, et que de la même
manière, 1/3 des plantes à
fleurs mauves sont “pures”
puisqu’elles sont homozy
gotes pour l’allèle A du gène.
d. je vérifie la validité de ma
théorie en vérifiant cette pré-
diction. En effet, je laisse ces
plantes de deuxième généra-
tion s’autoféconder et je
plante les graines ainsi obte-
nues pour avoir des plantes de
troisième génération.
Je constate que :
les plantes à fleurs blanches
étaient bien homozygotes aa
puisque leurs graines ne
donnent que des descen-
dants à fleurs blanches,
qu’un tiers des plantes à
fleurs mauves étaient bien
homozygotes AA, car leurs
graines ne donnent que des
descendants à fleurs mauves,
enfin que les deux autres
tiers étaient bien hétérozy-
gotes Aa puisque leurs
graines donnent pour 50 %
des plantes à fleurs mauves
et pour 50 % des plantes à
fleurs blanches.
Quelques précisions
d’histoire et de
vocabulaire
A l’époque, en 1865, les résultats de
Mendel ont été trouvés intéres-
sants, mais sans plus. Il fallut
attendre le début du siècle suivant
pour découvrir leur valeur universelle
et surtout pour comprendre que la
“loi de pureté” des gamètes,
postulée par Mendel, n’était pas un
postulat, mais la conséquence pure
et simple du comportement des
chromosomes qui portent les gènes.
En effet, tout individu reçoit un lot
de chromosomes de son père et
un autre lot de sa mère, de sorte
qu’il contient tout “en double”, deux
exemplaires de chacun des gènes,
un exemplaire d’origine paternelle et
un autre exemplaire d’origine
maternelle.
Puis tout individu, comme parent,
ne transmet que la moitié de ses
chromosomes, donc un seul exem-
plaire de chacun des gènes, de sorte
que son descendant aura, pour
chaque gène, un génotype, consti-
tué de deux exemplaires de ce gène,
un exemplaire venant de lui-même
et un autre exemplaire venant de
son conjoint.
Supposons un gène A qui comporte
plus de deux types différents d’al-
lèles : par exemple A1, A2 et A3.
Père et mère peuvent, pour un gène
donné, être hétérozygotes soit, par
exemple, A1/A2 et A1/A3. Puisque
chaque parent ne transmet qu’un
seul des allèles qui déterminent le
phénotype, ils peuvent engendrer
des enfants génétiquement diffé-
rents (A1A1 ou A1A2 ou A1A3 ou
A2A3), selon le hasard de la trans-
mission des allèles.
La conséquence de l’hérédité
mendélienne est que les parents ne
transmettent pas leurs caractères
mais seulement la moitié des gènes
qui déterminent, chez leurs enfants,
ces caractères. Pourquoi alors parle-
t-on de “caractères héréditaires” si
précisément ils ne le sont pas ?
Cela provient de la confusion sou-
vent entretenue entre les termes de
caractère et de phénotype.
On a vu qu’un caractère peut
apparaître sous plusieurs form e s
appelées phénotypes. On devrait
dire que la conséquence de l’hérédi-
té mendélienne est que les parents
ne transmettent pas leurs phéno-
types, mais seulement la moitié des
gènes qui déterminent, chez leurs
enfants, ces phénotypes.
Application des
principes mendéliens
aux maladies
héréditaires
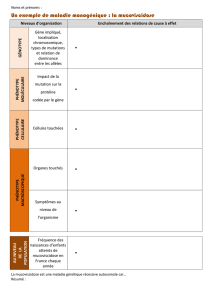
Les maladies mendéliennes
En application de ces principes, on
dit qu’une maladie est mendélienne
ou monogénique si, comme la diffé-
rence de couleur du pois, la différen-
ce entre les phénotypes sains et
malades ne dépend que d’un seul
gène.
Allèles normaux et allèles
pathologiques
Pour le gène impliqué dans une
maladie, on distingue le ou les allèles
dits “normaux” et le ou les allèles
Pour témoigner à Toulouse,
inscrivez-vous à :
AIRG - BP 78
75261 PARIS Cedex 06

11
NEPHROGÈNE N°34
Avril 2003
À LA GÉNÉTIQUE HUMAINE
À LA GÉNÉTIQUE HUMAINE
DU PETIT POIS DE MENDEL
DU PETIT POIS DE MENDEL
dits “pathogènes” ou “patholo-
giques” car leur présence est asso-
ciée à l’état pathologique de l’indivi-
du.
Les allèles pathogènes sont aussi
dits allèles mutés, et parfois plus
simplement “mutations” car ils
résultent d’une modification, c’est-à-
dire d’une mutation, d’un allèle
originellement normal. Une mutation
est une modification de la séquence
d’ADN du gène sous l’effet
d’éléments exogènes (radiations,
agents chimiques comme les gou-
drons du tabac) ou plus souvent
endogènes (radicaux libres) ou à
la suite d’erreurs de réplication
de l’ADN lors des divisions cellu-
laires.
Maladies autosomiques et
maladies liées au sexe 2
On dit qu’une maladie mendélienne
est autosomique si le gène impliqué
est porté par un autosome, c’est-à-
dire l’un des 22 chromosomes
humains, qui n’est ni le X, ni le Y.
Une maladie mendélienne est dite
liée au sexe si le gène impliqué est
porté par un chromosome sexuel,
soit X, soit Y, de sorte que la
transmission de la maladie dépend
de la transmission du sex e .
Par exemple, une maladie résultant
d’un gène porté par le chromosome
Y ne peut toucher que les garçons et
est transmise de père en fils.
Les maladies résultant des
mutations d’un gène porté par le
chromosome X seront vues plus
loin.
Maladies dominantes et
maladies récessives
On distingue les maladies domi-
nantes et les maladies récessives
comme on a distingué les phéno-
types dominants ou récessifs pour le
caractère de la couleur chez le pois.
Une maladie est dite dominante si la
présence d’un seul allèle patholo-
gique est suffisante, de la même
façon que le phénotype mauve de la
fleur de pois est dominant car la pré-
sence d’un seul exemplaire A du
gène est suffisante (AA et Aa sont
mauves).
Une maladie est récessive si les
deux exemplaires du gène consti-
tuant le génotype sont des allèles
pathologiques, de la même façon
que le phénotype blanc de la fleur de
pois est récessif car les deux exem-
plaires doivent être a pour que ce
phénotype soit réalisé.
Dans le cas d’une maladie récessive,
l’hétérozygote Aa est dit “porteur
s a i n” puisque son phénotype est
sain, mais qu’il est porteur d’un allè-
le pathologique (chez le pois l’hété-
rozygote Aa a des fleurs mauves,
mais est porteur de l’allèle a).
Comprendre pourquoi ou comment
une maladie est dominante alors
qu’une autre est récessive suppose
de connaître la fonction du gène et
les effets des diverses mutations qui
peuvent affecter celui-ci. Comme le
montre le schéma ci-dessus,
maladies dominantes et récessives
se distinguent par des modes de
transmission bien différents.
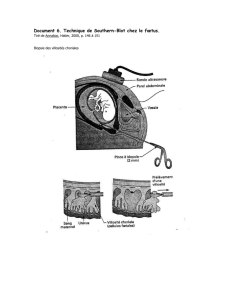
Arbres généalogiques.
Les hommes sont représentés par des carrés et les femmes par des cercles. Les
sujets atteints sont en noir. Les sujets décédés sont indiqués par trait diagonal.
Le sujet consultant est indiqué par une flèche.
A gauche, une famille avec une maladie récessive et à droite une famille avec une
maladie dominante.
Le livret Cystinose est accompagné par un supplément spécifique
pour les patients vivant en Suisse ou en Belgique.

12
NEPHROGÈNE N°34
Avril 2003
Mode de transmission d’une
maladie autosomique
dominante
Les généalogies des patients pré-
sentent les signes caractéristiques
suivants :
●
les deux sexes sont également
atteints,
●
tout individu atteint a l’un de ses
parents également atteint,
●
tout individu atteint transmet sa
maladie à la moitié de ses enfants,
●
tout individu indemne ne transmet
pas la maladie.
On observe donc une véritable
“transmission continue de la
maladie” au cours des générations.
Il est facile d’interpréter ce résultat
si on considère que :
●
le gène M impliqué dans cette
maladie est localisé sur un auto-
some ; ainsi les deux sexes ont
autant de risque d’être atteints,
●
que les individus atteints sont
hétérozygotes pour ce gène,
c’est-à-dire porteurs d’un allèle
n o rmal N (fonctionnel) et d’un
allèle muté M (pathogène), dont
l’effet est dominant sur celui de
l’allèle normal ; ainsi ils transmet-
tent bien leur exemplaire pathogè-
ne à la moitié de leurs descen-
dants qui sont atteints en raison
de la dominance de l’effet
pathogène de M ; leurs enfants
indemnes, ayant reçu un exem-
plaire normal N de chacun des
parents ne “transmettent” pas
la maladie (ce n’est pas la maladie
qui est transmise, mais l’allèle qui
en est la cause et qui, du fait de
son effet dominant, simule le fait
que la maladie est transmise !).
Mais il peut arriver qu’un sujet
atteint d’une maladie dominante
autosomique certaine naisse de
parents indemnes. Cette apparente
exception aux règles est due au fait
que dans un des gamètes paren-
taux, le gène normal a subi une
mutation générant un allèle patholo-
gique.
Mode de transmission d’une
maladie autosomique récessive
Une maladie “autosomique récessi-
ve” ou dont “le mode de transmis-
sion est autosomique récessif” se
caractérise par les observ a t i o n s
suivantes :
●
les deux sexes sont également
atteints,
●
tout individu atteint a deux
parents non atteints, formant un
couple à risque,
●
un couple à risque a, en moyenne
un quart de ces enfants atteints,
●
le risque de naissance d’un enfant
atteint est augmenté si les
parents sont apparentés.
Il est facile d’interpréter ce résultat
si on considère que la manifestation
de la maladie exige la présence de
deux exemplaires mutés et patho-
gènes, désignés par m, du gène
impliqué dans cette maladie.
Dans ce cas, chacun des deux
parents, qui n’est en général pas
atteint est dit “porteur sain” :
il est hétérozygote N/m, porteur de
l’allèle pathogène m qu’il a transmis
à son enfant atteint, et d’un allèle
“normal” fonctionnel N, dont l’effet
est dominant sur celui de l’allèle
pathogène. On dit aussi que l’effet
de l’allèle pathogène est récessif
vis-à-vis de l’effet de l’allèle normal.
Ce type de mode de transmission
concerne des maladies comme la
mucoviscidose, la phénylcétonurie
ou les thalassémies.
Maladies liées au sexe
Les maladies mendéliennes liées au
sexe sont principalement celles qui
sont associées à un gène du
chromosome X.
On sait que les femmes possèdent
deux chromosomes X alors que les
hommes possèdent un X et un Y. De
ce fait, les organismes masculins ne
possèdent qu’un exemplaire des
gènes du chromosome X : un allèle
pathogène du X induit toujours la
pathologie dans le sexe masculin.
Au contraire, dans le sexe féminin, il
y a deux exemplaires pour les gènes
du chromosome X et la pathologie
liée à l’X dépendra de la dominance
ou de la récessivité des effets des
allèles pathogènes. Cependant, ces
effets de dominance et de récessivi-
té interagissent avec le phénomène
d’inactivation de l’X. Cette inactiva-
tion est un phénomène complexe qui
fait que l’un des deux chromosomes
X est inactivé dans les cellules d’un
organisme féminin :
●
d’une cellule à l’autre, ce peut
être le chromosome X paternel
ou le chromosome X maternel
qui est inactivé ;
●
le pourcentage d’inactivation
du X paternel et du X maternel
est variable d’un tissu à
l’autre.
Les maladies monogéniques “réces-
sives liées au sexe”, comme l’hémo-
philie (qui a atteint la descendance
de la reine Victoria) ou la myopathie
À LA GÉNÉTIQUE HUMAINE
À LA GÉNÉTIQUE HUMAINE
DU PETIT POIS DE MENDEL
DU PETIT POIS DE MENDEL
 6
7
8
6
7
8
1
/
8
100%