Cycle cellulaire


Introduction
•Théorie cellulaire : toute cellule est issue d’une cellule
•L’information génétique est transmise aux 2 cellules filles à chaque
division
•Le génome de la cellule eucaryote :
–le matériel génétique est enfermé dans un compartiment membranaire : le
noyau
–matériel génétique = chromosomes
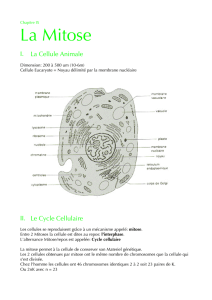
Rappel sur les chromosomes
•Chez l’homme : 23 paires soit 46 chromosomes contenant chacun 1 molécule
d’ADN
•Ne pas confondre brins d’ADN et molécule d’ADN !
1 chromatide = 1 molécule d’ADN = 2 brins d’ADN
1 molécule d’ADN = 1 fibre nucléosomique
•1 chromosome peut contenir 1 ou 2 chromatide(s) selon le moment du cycle cellulaire
•1 chromatide est formée d’1 molécule d’ADN
•1 molécule d’ADN est formée de 2 brins complémentaires

Vue d’ensemble du cycle cellulaire
•Il dure environ 10h chez l’Homme
•La mitose dure environ 1h chez l’Homme

•La phase G2 = transition fin réplication/mitose
→ synthèse d’ARNm et de protéines
•La mitose = division nucléaire (caryocinèse) puis division cytoplasmique
(cytodiérèse)
→ 1 cellule mère → 2 cellules filles
→ pas ou peu de synthèse d’ARNm et de protéines
Quand la cellule est en G1, si les conditions deviennent défavorables, retour
possible en G0 mais la passage de G1 à S est irréversible.
G0 G1 S
•La phase G0 = phase de repos
→ attente de conditions favorables pour
entrer dans un cycle cellulaire
•La phase G1 = accroissement de la cellule
→ synthèse d’ARNm et de protéines
•La phase S = réplication de l’ADN
(quantité ×2)
→ 1 molécule d’ADN → 2 molécules d’ADN
→ synthèse d’ARNm et de protéines

Le contrôle du cycle cellulaire
Il en existe 3 :
•G1→ S :
environnement favorable?
taille de la cellule (×2)?
•G2→ M :
ADN répliqué?
ADN intact?
•Métaphase→ Anaphase :
chromosomes alignés dans le plan équatorial?
chromosomes attachés au fuseau mitotique?
1) Points de contrôle
Le franchissement de ces 3 points de contrôle est sous le contrôle des
complexes protéiques Cdk = Cyclines dépendantes des Kinases.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%