Dossier_B-FR-03-02

NOTIFICATION DE DISSÉMINATION DE
PLANTES SUPÉRIEURES GÉNÉTIQUEMENT MODIFIÉES
(ANGIOSPERMAE ET GYMNOSPERMAE)
Directive 2001/18/CE
Essai au champ de maïs résistant aux insectes
Evénement 3243M
SYNGENTA SEEDS S.A.S
Janvier 2003
Syngenta Seeds SAS 1/28

PREAMBULE
La mise au champ de plantes génétiquement modifiées est soumise à une réglementation
européenne décrite dans la directive européenne 90/220 transposée dans la loi française du 13
juillet 1992. Cette directive 90/220 vient par ailleurs d’être modifiée et remplacée par la directive
2001/18 du 17 avril 2001. Cette nouvelle directive conforte le principe d’une progression par étape
qui permet d’adapter progressivement l’ampleur de la dissémination en fonction des
connaissances acquises sur ces plantes et notamment sur leurs effets sur la santé humaine et
l’environnement. Ainsi, en fonction de l’état d’avancement des dossiers au sein du processus
réglementaire (de la recherche amont vers le développement et la mise sur le marché) le niveau
des informations acquises et disponibles sera différent et beaucoup plus élevé pour un événement
de transformation en cours d’évaluation dans le cadre d’une autorisation de mise sur le marché
(utilisation à grande échelle) que pour un essai de type recherche (culture très localisée pendant
une courte période).
Ce dossier concerne une demande d’autorisation pour réaliser des essais permettant de
collecter du matériel végétal qui sera utilisé pour conduire des analyses de types
réglementaires, sur le matériel génétiquement modifié en comparaison avec du matériel non
génétiquement modifié.
Syngenta Seeds SAS 2/28

SOMMAIRE
INTRODUCTION
DOSSIER TECHNIQUE :
A. INFORMATIONS GÉNÉRALES
B. CARACTÉRISTIQUE BIOLOGIQUE DE L’ESPÈCE VÉGÉTALE RÉCEPTRICE
C. INFORMATIONS SUR LA MODIFICATION GÉNÉTIQUE
D. INFORMATIONS CONCERNANT LA PLANTE SUPÉRIEURE
GÉNÉTIQUEMENT MODIFIÉE
E. INFORMATIONS CONCERNANT LE SITE DE DISSEMINATION
F. INFORMATIONS CONCERNANT LA DISSÉMINATION
G. INFORMATIONS SUR LES PLANS DE SURVEILLANCE, DE CONTRÔLE,
ET DE TRAITEMENT DU SITE ET DES DÉCHETS APRES DISSÉMINATION
EVALUATION DES RISQUES POUR L’ENVIRONNEMENT
Syngenta Seeds SAS 3/28

INTRODUCTION
Les plantes sont soumises, au cours de leur culture, à des attaques de différents
pathogènes ( champignons, virus, insectes….) qui conduisent à des pertes de rendement et
des dépréciations de qualité.
Syngenta Seeds développe des programmes de recherche visant à réduire l’impact de ces
pathogènes sur les plantes cultivées. Parmi eux, la pyrale qui est un insecte lépidoptère
cause des dommages sur les plantes et les épis de maïs provoquant des baisses de
rendement et une plus grande sensibilité des plantes à d’autres attaques parasitaires.
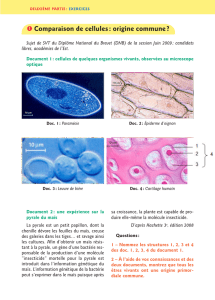
Le maïs
Le maïs (Zea mays) est une céréale annuelle originaire d’Amérique centrale qui est
maintenant cultivée du nord de l’ Europe au sud de l’Argentine , du niveau de la mer à des
altitudes supérieure à 300 mètres. Sous nos climats, elle est semée entre la mi-mai et la mi-
avril pour être récoltée en octobre.
Cette plante est cultivée sur 130 millions d’hectares dans le monde avec une production
annuelle d’environ 590 millions de tonnes. C’est la première production céréalière mondiale
essentiellement fournie par les Etats-Unis (230 millions de tonnes) suivi par la Chine, l’Union
Européenne (39 millions de tonnes), le Brésil et le Mexique. La France assure près de la
moitié de la production européenne avec une surface cultivée d’environ 1 800 000 hectares,
principalement dans les régions Aquitaine et Midi-Pyrénées. A coté de cette production de
maïs grain, il faut également signaler, une production de maïs fourrage, dans les zones
Ouest et Nord de la France.
Le maïs grain est essentiellement utilisé en alimentation animale et pour une part moins
importante à l’industrie de l’amidon et à la semoulerie.
Les attaques de pyrale
La pyrale est une espèce d’insecte lépidoptère présente sur la majeure partie du territoire
français.
Le cycle évolutif de la pyrale du maïs, Ostrinia nubilalis, passe par quatre stades : œuf,
larve (chenille), nymphe (chrysalide) et papillon.
On observe une seule génération par an dans le Nord (cycle univoltin) et deux générations
(cycle bivoltin) dans le Midi méditerranéen. Dans les autres régions, selon les conditions
climatiques, un certain nombre seulement de larves de première génération poursuit son
développement pour donner des individus de deuxième génération.
L’adulte de la pyrale est un papillon de 2 à 3 cm d’envergure qui,
l’été, dépose ses œufs (ooplaques d’une vingtaine d’unités) à la
face inférieure des feuilles de plantes de maïs situées au-dessus
ou en-dessous de l’épi. Après une incubation dont la durée varie
de 1 à 4 semaines en fonction de la température, les larves de
pyrale apparaissent et se « baladent » sur les feuilles et la tige
(stade baladeur) puis elles pénètrent dans la plante par la partie
sommitale de la plante, le cornet.
Syngenta Seeds SAS 4/28

Elles s’alimentent en réalisant des perforations, caractéristiques d’une attaque précoce, sur
les feuilles encore repliées. Elles s’installent ensuite dans l’axe central du panicule mâle,
provoquant ainsi des casses de panicules. Puis elles s’établissent dans la tige et dans l’épi
lui-même creusant des galeries où elles ne peuvent être traitées par des moyens biologiques
et chimiques classiques.
(source INRA)
Elles mangent la tige et détruisent les vaisseaux responsables du transport des éléments
nutritifs de la plante. A l’arrivée des premiers froids les larves redescendent à la base de la
tige pour passer l’hiver en diapause. Avec l’arrivée des jours longs et des températures plus
élevées, les larves hibernantes se transforment en nymphes pendant les mois de mai-juin.
Les papillons de première génération émergeront, sur une période d’un mois environ, en
juin-juillet.
Cet insecte ravageur provoque différents types de dégâts sur les cultures de maïs :
- La casse des panicules mâles au niveau de la dernière feuille ce qui limite l’émission de
pollen et pénalise la formation de grains. (source INRA)
- La casse des plantes dues aux galeries forées dans la tige. Les galeries dans la rafle de
l’épi entraînent la chute des épis.
- La recrudescence des pathogènes dont les galeries servent de porte d’entré. La perforation
des épis favorise ainsi le développement de champignons pathogènes, les Fusarium, qui
produisent des mycotoxines susceptibles d’être dangereuses pour l’homme et les animaux.
Même sans dégâts apparents, la présence de la chenille entraîne un affaiblissement de la
plante se traduisant par une diminution du poids unitaire du grain. Le seuil de nuisibilité est
estimé à une chenille par plante à la récolte pour les cultures de maïs grain.
Dans certaines régions, les destructions peuvent représenter jusqu’à 30 % de la récolte. Au
plan mondial, 7 % environ de la récolte annuelle est endommagée par cet insecte. En
France on estime que 70% des surfaces de maïs grain sont infestées, causant près de 50
millions d'euros de pertes.
Syngenta Seeds SAS 5/28
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
1
/
28
100%