Physiologie Rénale : Résumé de Cours

Cours de physiologie rénale
Résumé :
introduction :
les organes du système urinaire sont les reins, les uretères, la vessie et l’urètre.
L'eau et les solutés qui restent après les reins ont filtré le sang est retourné la majeure partie
de l'eau et des solutés à la circulation sanguine constituent l'urine.
Fonctions du rein: vue d'ensemble
les reins régulent la composition ionique du sang, son osmolarité, le volume sanguin, la
pression artérielle ainsi que le pH du sang.
Les reins participent aussi à la néoglucogenèse, libère le calcitriol et l'érythropoïétine et
excrètent les déchets et les substances étrangères.
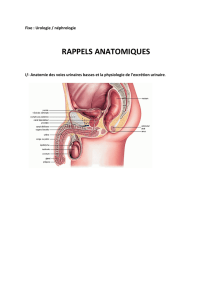
Anatomie et histologie des reins.
Les reins sont des organes rétro péritonéaux fixés à la paroi abdominale postérieure.
Trois couches de tissu enveloppent les reins : la capsule fibreuse, la capsule adipeuse, et le
fascia rénal.
L'intérieur du Rhin est formé d'un cortex, d'une médullaire, de pyramides, de papilles et de
colonnes rénales, de calices et d’un bassinet.
Le sang pénètre par l'artère rénale. Il passe, dans l'ordre, par les artères segmentaires,
interlobaires, arquées et interlobulaires. Les artérioles glomérulaires efférentes, les capillaires
péritubulaires, les artérioles et les veinules droites (aussi appelés collectivement vasa recta), et
les veines interlobulaires, arquées, interlobaires, et segmentaire. Enfin il quitte le rein par la
veine rénale.
Des nerfs vasomoteurs de la partie sympathique du système nerveux autonome innervent les
vaisseaux sanguins du rein. Il participe à la régulation du débit sanguin.
Le néphron est l'unité fonctionnelle du rein. Il est constitué d'un corpuscule rénal (glomérule
et capsule glomérulaire ou de bowman) et d'un tubule rénal.
Le tubule rénal comprend le tubule contourné proximal, l’anse du néphron est formée d'une
partie descendante et d'une partie ascendante.
Le néphron cortical à une anse courte qui ne pénètre pas la région superficielle de la
médullaire rénale. : Le néphron juxta médullaire à une anse longue qui s'enfonce dans la
médullaire presque à la papille rénale.
La paroi de la capsule glomérulaire, du tubule rénal et du tubule collecteur est formée, d'un
bout à l'autre d'une seule couche de cellules épithéliales. L'épithélium de chaque partie du
tubule possède des caractéristiques histologiques du tubule rénal et du tubule collecteur.
L'appareil Juxta glomérulaire est composé de cellules Juxta glomérulaires et de la macula
densa de l'extrémité de la partie ascendante de l’anse du néphron.
Physiologie rénale : vue d'ensemble
Le néphron assure trois fonctions principales : la filtration glomérulaire, la sécrétion tubulaire,
et la réabsorption tubulaire.
Filtration glomérulaire

Le liquide qui pénètre dans la chambre glomérulaire est appelé filtrât glomérulaire.
La membrane de filtration endothélio capsulaire est formée de l'endothélium glomérulaire de
la membrane basale et défendent filtration des pédicelles des podocytes.
La plupart des substances du plasma traversent facilement le filtre glomérulaire. Toutefois les
globules sanguins et la plupart des protéines ne passent pas dans le filtre.
Le filtre glomérulaire totalise jusqu'à 180 l de liquide par jour. La quantité de fluide filtré est
élevée parce que le filtre est mince et pour eux, les capillaires glomérulaires sont longs et la
pression sanguine dans les capillaires est élevée.
La pression hydrostatique glomérulaire favorise infiltration, alors que la pression
hydrostatique capsulaire et la pression oncotique s'y oppose. La pression nette de filtration et
d'environ 10 mm de mercure.
Le débit de filtration glomérulaire et la quantité de filtra produit dans les deux rats par minute
: il est normalement 105 à 125 ml par minute.
Le débit de filtration glomérulaire dépend de l'autorégulation rénale, de la régulation nerveuse
et de la régulation hormonale.
Réabsorption et sécrétion tubulaires.
La réabsorption tubulaire est un processus sélectif qui récupère les substances du fluide
tubulaire et les retourne à la circulation sanguine. Les substances réabsorbées comprennent
l'eau le glucose les acides aminés et des ions tels que le sodium le chlorure le potassium le
bicarbonate et le phosphate.
Certaines substances dont l'organisme n'a pas besoin sont retirés de la circulation sanguine et
rejeté dans l'urine par sécrétion tubulaire. Ce sont entre autres, des ions K+, H+ NH4+, l'urée,
la créatinine et certains médicaments.
La réabsorption s'effectue par les voies paracellulaire (entre les cellules et les tubules) et
transcellulaire (à travers les cellules des tubules).
La quantité maximale d'une substance qui peut être réabsorbée par unité de temps s'appelle le
transport maximal(Tm).
Environ 90 % de la réabsorption d'eau est obligatoire : elle s'effectue par osmose et
accompagne la réabsorption des solutés. Les 10 % qui restent constituent la réabsorption
facultative de l'eau, qui varie selon les besoins de l'organisme et obéit à la régulation de
L’ADH ou hormone anti diurétique.
Les ions sodium sont absorbé sur toute l'étendue de la membrane basale latérale par transport
actif primaire.
Dans le tubule contourné proximal, les ions sodium sont réabsorbés à travers la membrane
apicale par les symporteurs sodium/glucose et les anti porteurs sodium/H+ ; l'eau est
réabsorbée par osmose, les ions chlorure, potassium, calcium et magnésium ainsi que l'urée
sont réabsorbées par diffusion passive ; l'ammoniaque et l'ammonium sont sécrétées.
L’anse du néphron réabsorbe de 20 à 30 % des ions sodium et potassium ainsi que calcium et
hydrogèno-carbonate, 35 % des ions chlorure et 15 % de l'eau filtrée.
Le tubule contourné distale absorbe les ions sodium et chlorure au moyen d'un symport
sodium/chlorure.
Dans le tubule collecteur, les cellules principales réassort des ions sodium et ses graines des
ions potassium. Certaines cellules intercalaires ré-absorbent des ions potassium et hydrogèno-

carbonate et sécrète des ions H + alors que d'autres cellules intercalaires sécrètent des ions
hydrogèno- carbonate.
L'angiotensine II, l'aldostérone, l'hormone antidiurétique et le peptide natriuretique auriculaire
régule la réabsorption des solutés et de l'eau.
En l'absence d'ADH, les reins produisent de l'urine diluée, les tubules rénaux absorbent plus
de soluté que d'eau.
En présence d'ADH, les reins produisent de l'urine concentrée, de grandes quantités d'eau sont
réabsorbées du fluide tubulaire et passe dans le liquide interstitiel, ce qui augmente la
concentration des solutés dans l'urine.
Le mécanisme à contre-courant établit un gradient osmotique dans le liquide interstitiel de la
médullaire rénale, qui permet la production d'urine concentrée lorsque l'ADH est présente.
Évaluation de la fonction rénale
l'examen des urines est une analyse du volume des propriétés chimiques, physiques et
microscopiques d'un échantillon d'urine.
Sur le plan chimique l'urine normale contient 95 % d’eau et 5 % de soluté. Parmi les solutés
normalement présents se trouve l’urée, la créatinine, l'acide urique, l’urobilinogène et divers
ions.
Des constituants anormaux qu'un examen des urines peut révéler, dont l'albumine, le glucose,
les globules rouges et blancs, les corps cétoniques, la bilirubine, l’urobilinogène en quantité
excessive, les cylindres urinaires et les microbes.
La clairance rénale est la capacité des reins à retirer une substance donnée du sang par unité
de temps.
Transport, entreposage et élimination de l'urine.
Les uretères sont rétro péritonéaux et sont constitués d'une muqueuse, d'une musculeuse et
d'une adventice. Il transporte l'urine du bassinet à la vessie principalement par péristaltisme.
La vessie est située dans la cavité pelvienne derrière la symphyse pubienne ; sa fonction est
d’entreposer l'urine entre les mictions.
La vessie est composée d'une muqueuse avec des replis muqueux, d'une musculeuse (muscle
vésical) et d'une adventice (céreuse sur la face supérieure).
Le réflexe de la miction évacue l'urine de la vessie par suite d'influx parasympathiques qui
causent la contraction du muscle vésical et le relâchement du sphincter lisse de l'urètre et par
suite de l'inhibition des influx dans les neurones moteurs somatiques qui innervent le muscle
sphincter de l'urètre.
Murray est un conduit qui commence dans le plancher de la vessie jusqu'à l'extérieur du
corps. Son anatomie et son histologie sont différents chez l'homme et la femme. Chez les
deux sexes l'urètre sert à l'évacuation de l'urine du corps ; si la mission également à l'émission
du sperme.
Traitement des déchets ailleurs dans l'organisme
Outre les reins, plusieurs tissus, organes et processus contribue au confinement temporaire des
déchets, à leur transport en vue de l'évacuation, au recyclage des matériaux et à l'expression
des substances excédentaires toxiques.

Les tampons lient les ions H+ excédentaire, le sang transporte les déchets, le foie convertit les
substances toxiques en substance moins toxiques, les poumons dégagent du CO2, les glandes
sudoripares contribuées à dissiper la chaleur et le tube digestif élimine les déchets solides.
Cours
introduction :
Le rein joue un rôle capital dans la régulation de la pression artérielle et dans l’homéostasie ;
il participe à l'élimination des substances (urée) et toxines. Les maladies rénales et du tractus
urinaire sont dues aux désordres hydroélectrolytiques comme la rétention de sels,
l'augmentation du volume plasmatique, l'œdème, l'insuffisance cardiaque, l'acidose et
hyperkaliémie.
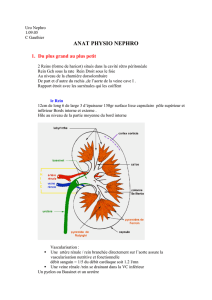
Anatomie du rein.
Les reins ont une forme de haricot, d'environ 12 cm de haut. La face interne est convexe. Il y
a le hile dans le creux du rein qui permet la circulation sanguine. Les reins sont situés dans la
zone lombaire. Le rein droit est situé plus bas que le gauche.
Les reins sont entourés d'une capsule qui lui donne un aspect lisse. Il y a au-dessus du rein un
tissu graisseux ;le fascia rénal ,et la graisse para rénale. Le Fascia rénal permet la fixation
dans l’abdomène. Sur une coupe du rein on observe plusieurs structures. Le cortex se
prolonge dans les pyramides médullaires. La médullaire est striée et chaque pyramide se
dirige vers le hile. il y a huit lobes rénaux. Chaque pyramide rénale va dans une papille puis
un calice mineur puis un calice majeur, dans le bassinet pour enfin finir dans l'uretère. Une
onde péristaltique est possible pour permettre le passage de l'urine dans la vessie.
Vascularisation :
le rein ne représente que 0,5 % du poids du corps mais 1200 ml de sang y passe par minute.
Les artères interlobaires cheminent jusqu'à la base de la pyramide pour donner des artères
arquées. Le sang ressort par les veinules en suivant le chemin inverse.
Innervation sympathique
les néphrons
chaque rein en contient 1 million. C'est un tube recouvert d'épithélium monocouche à
l'intérieur. Il est divisé en plusieurs parties. Les globules se prolongent par un tube ; et sont
situés dans le cortex du rein. Il y a un réseau vasculaire dans chacune. L'endothélium repose
sur une membrane basale. Les podocytes reposent sur la membrane basale. Les porzs se
situent entre deux pédicelles. L'albumine ne passe pas dans l'urine.
Le tubule contourné proximal.
Les bronches ascendantes et descendantes forment le tube de henle. Chaque segment du
tubule à une fonction propre. La anse de Henle à un épithélium plat. Les cellules du tubule
contourné contiennent une bordure en brosse. La médullaire est essentiellement constituée de
tubes. L'artère glomérulaire afférente à une pression de 60 mm de mercure. Cette pression
diminue un peu dans les capillaires. Les capillaires et tubulaires rejoignent le système cave.
Les capillaires Vasa recta suivent la anse de henle. Les capillaires offrent peu résistance, il
n'y a pas de changements de pression. La pression élevée est un moteur de la filtration. Le
filtrat glomérulaire atteint 180 l par jour. Tous les globules sont dans le cortex (85 %).

L’appareil juxtaglomérulaire est à la jonction des glomérules et de l'épithélium. Il y a une
différenciation des cellules qui donnent la macula densa.
Les processus de base de la physiologie rénale.
La filtration glomérulaire.
Au cours de la première étape de la production de l'urine une partie de l'eau et la plupart des
solutés plasma quittent la circulation sanguine en traversant la paroi des capillaires
glomérulaires et passe dans la capsule glomérulaire, qui se jette dans le tubule rénal.
La réabsorption tubulaire.
Lorsque le filtrat s'écoule dans le tubule rénal et le tubule rénal collecteur, environ 99 % de
l'eau et un grand nombre de solutés utiles sont réabsorbés par les cellules des tubules et
regagne le sang qui circule dans les capillaires péritubulaires et les artérioles ainsi que
veinules droites. Notez que le terme réabsorption désigne le retour de substances à la
circulation sanguine, alors que le terme absorption désigne l'entrée de nouvelles substances
dans l'organisme comme cela se produit dans le tube digestif.
La sécrétion tubulaire.
Au fur et à mesure que le liquide passe dans le tubule rénal et le tubule rénal collecteur, les
cellules de ces conduits y sécrètent des substances additionnelles telles des déchets, des
médicaments et des ions excédentaires. Notez que la sécrétion tubulaire retire des éléments du
sang, dans les autres cas de sécrétion, comme la sécrétion d'hormones ; les cellules libèrent
une substance dans le liquide interstitiel et le sang.
Les solutés du liquide qui se déversent dans le bassinet restent dans l'urine et sont excrétés. Le
taux d’excrètion urinaire d'un soluté est égal à son taux de filtration glomérulaire + son taux
de sécrétion – son taux de réabsorption.
La filtration glomérulaire.
Le liquide qui pénètre dans la chambre glomérulaire est appelé filtrat glomérulaire. La
fraction du plasma qui quitte les artérioles glomérulaires afférentes des reins pour former le
filtrat glomérulaire est la fraction filtrée. En général la fraction filtrée est de 16 à 20 %, mais
cette valeur varie considérablement, chez les individus sains comme chez les malades. En
moyenne, le volume quotidien de filtrat glomérulaire est de 150 à 180 l. Toutefois plus de 99
% du filtrat glomérulaire retourne à la circulation sanguine par réabsorption tubulaire si bien
que le rein n'excréte quotidiennement qu'1 à 2 l d'urine par jour.
La membrane de filtration.
Elle est constituée de l’ensemble les cellules endothéliales des capillaires glomérulaires et des
podocytes qui entourent complètement les capillaires, forment une barrière poreuse appelée
membrane de filtration, ou membrane endothélio-capillaire. Cette disposition en forme de
sandwiches permet le passage de l'eau et des petits solutés mais retient la plupart des protéines
plasmatiques de même que les érythrocytes, les leucocytes et les plaquettes. Les substances
extraites du sang par filtration traversent trois barrières :
– une cellule endothéliale glomérulaire
– la membrane basale
– une fente de filtration formée par un podocyte.
Les cellules endothéliales des capillaires glomérulaires sont assez poreuse (grand pores) qui
mesurent de 0,07 à 0,1 µm de diamètre. La taille de ces ouvertures permet à tous les solutés
du plasma sanguin de quitter les capillaires glomérulaires mais s'oppose au passage des
érythrocytes, des leucocytes et des plaquettes. Parmi les capillaires glomérulaires et dans le
sillon entre les artérioles glomérulaires afférentes et efférentes se trouvent des cellules
mésangiales. Ce sont des cellules contractiles qui participent à la régulation de la filtration
glomérulaire.
La membrane basale qui est une couche de matière acellulaire, située entre l'endothélium et
les podocytes, est constituée de minuscules fibres de collagène et de protéoglycanes
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%