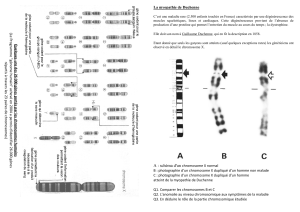
Cours génétique Mutations Boukhatem

CHAPITRE 3 : LES MUTATIONS
77
CHAPITRE 3
LES MUTATIONS
1. Introduction
1.1. Définition
Les mutations sont des changements brusques et d’emblée héréditaires de
l'information génétique. L'information héréditaire est normalement stable, elle est transmise
de génération en génération de manière fidèle. Mais cette fidélité n'est pas absolue. Le
système à la base de la transmission de l'information héréditaire est ce qu'on appelle la
réplication. La réplication permet de faire à partir d'une molécule ADN une copie conforme
(exacte). Cette copie est presque toujours conforme à la molécule originale, de sorte que le
génotype reste constant de la fécondation jusqu’à la mort. Mais il arrive que la réplication soit
accompagnée de quelques erreurs très rares aboutissant à des copies qui diffèrent du modèle
original: ce sont les mutations géniques.
En plus des mutations produites à la suite d'erreurs au niveau de la réplication, il y a
différents agents de l’environnement qui peuvent altérer la structure de la molécule d'ADN et
qui provoquent donc l'apparition des mutations. Ces agents peuvent êtres de différentes
natures (physique, chimique ou même biologique). Ce sont les agents mutagènes.
Le mot mutation est souvent employé dans un sens large pour designer non seulement
les gènes ayant subi une altération interne, mais aussi les variations structurales des
chromosomes. On distingue alors les deux catégories de mutations en qualifiant les premières
de géniques et les secondes de chromosomiques. En plus de ces deux types de mutations, il
existe une troisième catégorie dite mutation génomique où l'altération touche le nombre des
chromosomes (variation numérique des chromosomes).

CHAPITRE 3 : LES MUTATIONS
78
1.2. Mutations et phénotype
Tous les caractères héréditaires peuvent être touchés par les mutations. Ainsi, il y a
plusieurs sortes de mutations: morphologiques, physiologiques, biochimiques ou même
psychologiques. Chez la drosophile par exemple, il existe des mutations qui touchent: la
pigmentation des yeux ou du corps, les ailes, la répartition des soies, les caractères sexuels, la
fertilité, la longévité, les propriétés sérologiques, la réaction à la lumière, le comportement
durant l’accouplement... etc.
Les conséquences des mutations sur le phénotype varient des mutations dont l'effet sur
le phénotype est imperceptible aux mutations dont l'effet est dramatique (modification
radicale du phénotype).
Les mutations peuvent toucher tous les types de cellules d'un organisme. Chez les
métazoaires à reproduction sexuée, les mutations peuvent toucher soit des cellules somatiques
soit des cellules germinales. Les mutations touchant les cellules somatiques sont transmises
aux cellules filles (par suite de mitose), il en résulte qu'une partie des cellules expriment un
phénotype mutant, ce qui donne les chimères ou mosaïques. Les mutations somatiques ne
vont pas êtres transmis à la descendance et vont donc disparaître au plus tard avec la mort de
l’individu où ils sont apparues. Mais les mutations qui touchent une cellule germinale peuvent
être transmises à la descendance. Ces dernières sont les seules à pouvoir changer l’hérédité
dans une lignée ou une famille.
Dans les tissus haploïdes les effets des mutations sur le phénotype peuvent être décelés
dès l'apparition des mutations. Mais chez les diploïdes les mutations ne peuvent être décelées
dans la descendance immédiate de l’individu où elles sont apparues que si elles sont
dominantes. Si elles sont récessives, elles ne seront décelées que quelques générations après
leurs productions. Chez les polyploïdes les mutations sont décelées encore plus tardivement
que chez les diploïdes.

CHAPITRE 3 : LES MUTATIONS
79
1.3. Fréquences
Si on considère les fréquences des mutations au niveau d'un gène donné on constate
que les fréquences sont généralement très basses. Cependant ces fréquences varient
énormément d'un gène à un autre un sein d'une même espèce. Chez le Maïs par exemple,
Stadler n'a pas observé une seule mutation du gène (Wx) en (wx) parmi 1503744
gamètes testés, alors que parmi 554786 gamètes il a observé 273 mutations du gène (R) en (r)
.
Mais si le taux de mutation des gènes est assez bas, le taux global de l'ensemble des
gènes est assez important. Si on prend l'exemple de Drosophila melanogaster, au moins 5 %
des gamètes contiennent une nouvelle mutation à chaque génération. De la même façon que
les fréquences des mutations varient énormément d'un gène à un autre au sein d'une même
espèce, la fréquence globale des mutations par espèce varient énormément d'une espèce à une
autre.
2. Nature des mutations
2.1. Mutations ponctuelles
Ce sont des modifications qui touchent une seule paire de base à la fois ou un nombre
très faible de paires de bases. Ce sont donc des mutations géniques. Il y'a deux types de
mutations ponctuelles: les substitutions et les délétions/additions.
2.1.1. Substitutions
C'est le changement d'une paire de base par une autre. Au niveau de la molécule
d'ADN quatre types de bases sont présentes:
- La cytosine et la thymine qui sont des pyrimidines
- La Guanine et l’Adénine qui sont des purines.
Quand la substitution consiste en le changement de purines par des purines (exemple
A/G ou G/A) ou le changement d’une pyrimidine par une pyrimidine (T/C ou C/T), on dit que
c'est une transition. Quand la substitution consiste en le changement d'une purine par une

CHAPITRE 3 : LES MUTATIONS
80
pyrimidique (G/T ou A/C...) ou inversement changement d'une pyrimidine par une purine
(T/A ou T/G...), on dit que c'est une transvertion (figure 3.1).
Les substitutions de bases conduisent à des mutations qui peuvent avoir des
conséquences différentes au niveau du phénotype. On distingue trois types de mutations par
substitutions selon leur conséquence au niveau de la protéine et donc au niveau du phénotype.
Mutation silencieuse: le changement au niveau du génome est imperceptible au niveau du
phénotype car le changement de l’aminoacide, induit par la substitution au niveau de la
protéine, n'a pas d'effet sur l'activité de cette dernière. L'absence du changement au niveau du
phénotype peut être due au fait que le codon initial et le codon mutant codent tous les deux
pour le même aminoacide (figure 3.2).
Figure 3.1. Les différents types de mutations par substitution.

CHAPITRE 3 : LES MUTATIONS
81
séquence génique séquence ARNm
séquence protéique
Sauvage
AAC UUG Lys
Mutant
AAT UUA Lys
Figure 3. 2. Mutation silencieuse: changement de l'information au niveaux du génotype sans
effet sur le phénotype (le code génétique est dégénéré c'est à dire qu'un aminoacide peut être
codé par plusieurs codons différents). Lys: lysine.
Mutation faux-sens (miss-sens): le codon mutant spécifie un aminoacide diffèrent de celui
spécifié par le codon initiale. L'effet de ce changement au niveau du phénotype dépend de la
nature de l’aminoacide échangé, par rapport à celui d'origine (aminoacide de même groupe
chimique ou nom: aminoacide acide, basique ou neutre), et de l'emplacement de l’aminoacide
échangé (cite actif de l'enzyme ou en dehors du cite actif). Selon la nature chimique de
l’aminoacide mutant et son emplacement au niveau de la protéine le résultat peut être une
enzyme mutante inactive, moins efficace ou thermosensible (figure 3.3).
Séquence génique Séquence ARNm
Séquence protéique
Sauvage
CAA GUU Val
Mutant
GAA CUU Leu
Figure 3. 3. Mutation faux-sens (miss-sens). La mutation a pour conséquence le changement
d’un aminoacide par un autre au niveau de la protéine codée par le gène.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
1
/
32
100%