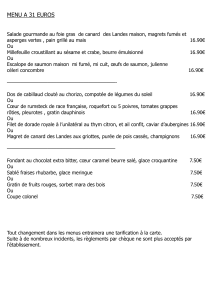
Role du nerf et du territoire cutane dans le

/.
Embryo!, exp. Morph. Vol. 27, 2, pp. 277-300, 1972 277
Printed
in
Great Britain
Role
du
nerf
et du
territoire
cutane dans
le
developpement
des
corpuscules
de
Herbst
et de
Grandry
Par RAYMOND SAXOD1
Labor atoire de Zoologie de VUniversite Scientifique et Medicate
de
Grenoble
(Directeur: Professeur P. Sengel)
SUMMARY
Role of nerve and
cutaneous
tissue in the development ofHerbsfs and
Grandry]s
corpuscles
Frontal buds
or
pieces
of
the bill
of
duck
and
chick embryos have been explanted
on the
chorioallantoic membrane, transplanted
as
homo-
or
xeno-plastic grafts
to the
flank,
the
frontal
bud and the
limb
bud of
early host embryos,
or
associated
as
coelomic grafts with
various isolated nerve sources. Results show that:
1.
The onset
of the
histogenesis
of
Herbst's
and
Grandry's cutaneous sensory corpuscles
is entirely dependent
on
the presence
of
a nerve ending, irrespective
of
the stage
at
which the
graft
is
obtained.
The
nerve ending
is
also required
for the
maintenance
of the
structural
integrity
of the
previously differentiated corpuscles
and for
their subsequent development.
2.
Only somato-sensory nerve endings are
able to
ensure the development of
the
corpuscles;
central connexions
are not
required. Sympathetic
or
somato-motor fibres
are not
able
to
sustain the development
of
the corpuscles.
3.
The corpuscle type,
as
well as corpuscle distribution,
is in
conformity with the origin
of
the integument
in
which
it
develops
and is
determined
by
the specificity and regional quality
of the innervated dermal mesenchyme.
The
determination
of
the cutaneous territory occurs
at
a
very early stage (prior
to the 3rd
day
of
incubation
in the
frontal
bud of
the duck).
4.
Heterotopic
or
xenoplastic innervation
of the
graft between duck
and
chick does
not
alter the cutaneous specificity
of
the differentiation. However, corpuscles did not differentiate
when duck frontal buds were innervated by mouse spinal ganglia.
These results are discussed in connexion with what is known about peripheric sense organs
in vertebrates.
A
schematic model
is
proposed
for the
mechanism
of
the morphogenesis
of
the corpuscles.
INTRODUCTION
De nombreux travaux recents ont ete consacres aux corpuscules sensoriels
des Vertebres. La plupart d'entre eux ont ete realises chez l'adulte et ont
trait a l'ultrastructure des differentes categories cellulaires qui les constituent
(voir la revue que nous en avons faite, Saxod, 1969).
Les etudes experimentales (revues de Wright, 1951; Singer, 1960; Zelena,
1 Adresse
de
Vauteur: Laboratoire
de
Zoologie, Universite Scientifique
et
Medicale
de
Grenoble, Domaine Universitaire, 38-Saint Martin-d'Heres, France.
l8
EHB 27

278 R. SAXOD
1964;
Hughes, 1968; Guth,
1969 a)
ont ete realisees principalement chez les
Poissons (Olmsted, 1920; Brockelbank, 1925; Parker, 1932; Bailey, 1937;
Kamrin & Singer, 1953; Denizot & Baillet-Derbin, 1969; voir aussi la revue de
Devillers, 1958), les Batraciens (Stone, 1922, 1933; Whiteside, 1927; Bedell,
1939;
Speidel, 1947, 1948, 1964; Wright, 1951, 1955, 1964; Sidman & Singer,
1960;
Poritsky & Singer, 1963; Robbins, 1967; Jones & Singer, 1969), et chez
les Mammiferes (May, 1926; Torrey, 1934, 1940; Wagner, 1953; Guth, 1958;
Kitamura, 1965; Oakley & Benjamin, 1966; Oakley, 1967; Farbman, 1969;
Zalewski, 1969; Olivieri-Sangiacomo, 1970; Iwayama, 1970; Fujimoto &
Murray, 1970).
Quant aux organes terminaux des Oiseaux, peu d'etudes recentes leur ont ete
consacrees. La plupart des recherches experimentales sont anciennes et consis-
terent a sectionner ou a intoxiquer le nerf peripherique (Botezat, 1910; Boeke,
1926;
Klein, 1932; Quilliam & Amstrong, 1961) ou a effectuer des greffes
heterotopiques chez l'adulte (Dijkstra, 1933). De nombreuses hypotheses,
souvent contradictoires, ont ete emises quant a l'organisation et l'origine des
corpuscules (voir les revues que nous en avons faites (Saxod, 1970a, b).
Parallelement a des recherches en microscopie electronique sur la structure
et l'histogenese des corpuscules sensoriels cutanes de Herbst et de Grandry chez
le Canard (Saxod, 1969, 1970 a, b), et sur l'origine des differentes categories
cellulaires qui les constituent (Saxod, 1971), j'ai realise des travaux experi-
mentaux afin d'analyser les facteurs de leur morphogenese.
Dans ce memoire, qui constitue une premiere partie de ce travail, j'ai etudie
les conditions de leur differenciation chez le Poulet et le Canard afin de repondre
aux questions suivantes: la terminaison nerveuse est-elle indispensable au
developpement et au maintien des corpuscules? Si oui, quelles categories de
fibres nerveuses sont-elles capables d'assurer leur developpement? Le type de
corpuscule forme est-il determine par le nerf ou par le territoire innerve?
Les
corpuscules sensoriels
cutanes du Poulet et du Canard
II existe divers types d'organes terminaux encapsules chez le Poulet et le
Canard; ils different par leur structure, leur densite et leur mode de repartition
dans le tegument des deux especes.
Les
corpuscules
de Herbst
Nous en avons distingue deux types (Saxod, 1967).
Ceux du type 'bee de Canard', localises exclusivement dans le bee du Canard,
ou ils sont abondants, et dans la langue du Canard. Ils sont
ovoides,
de dimensions
constantes (160 x 100 /*m chez l'adulte), le bulbe interne comprend une vingtaine
de noyaux disposes en deux rangees symetriques par rapport a la terminaison
nerveuse, l'espace interne est nettement lamellaire (Fig. 5).
Ceux du type 'bee de Poulet' qui sont presents dans le bee du Poulet et dans le
derme des diverses regions cutanees du Poulet et du Canard, a l'exclusion du

Le developpement des corpuscules de Herbst et de Grandry 279
bee et de la langue du Canard. Ils sont allonges, de dimensions variables chez
l'adulte (allant de 25 x 12 a 300 x 110 /on), le bulbe interne des corpuscules de
grande taille comprend jusqu'a une cinquantaine de noyaux qui tendent a se
placer sur plusieurs rangees, l'espace interne est moins nettement structure que
dans le type 'bee de Canard' (Fig. 6).
Les
corpuscules
de Grandry
Us sont localises exclusivement sous l'epiderme du bee du Canard, ou ils
abondent, et dans la langue. Ils mesurent 50 x 30 /*m chez l'adulte.
Un autre type de corpuscule, appele corpuscule de Merkel, existe dans la
region du palais du bee de Poulet, entre les digitations de l'epithelium. Cet
organe terminal, dont la taille (20 x
8 jum)
est nettement inferieure a celle du
corpuscule de Grandry, a cependant une structure assez voisine de ce dernier
(Andersen & Nafstad, 1968).
Le developpement des corpuscules de Merkel, difficiles a identifier avec
certitude aux stades embryonnaires, n'a ete etudie qu'occasionnellement dans
ce travail. Nous avons done generalement retenu pour l'analyse des resultats,
les corpuscules de Herbst de types 'bee de Poulet' et 'bee de Canard', et les
corpuscules de Grandry.
MATERIEL ET METHODES
Les divers types de greffes ont ete realises chez le Poulet (Leghorn blanc) et
le Canard (Pekin). L'age des donneurs et des embryons notes est indique dans
les Tableaux 2-4. A la fin de l'experience, au moment de la fixation, l'age du
greffon (son age absolu) est exprime, en jours d'incubation, par la somme: age
du donneur au moment du prelevement du greffon + duree du sejour du greffon
sur l'hote. Les greffons, preleves sur des bourgeons frontaux ou des bees
d'embryons, sont coupes en cubes ayant 0,5 a
1
mm de cote environ et maintenus
en place, lors des greffes sur bourgeon frontal et sur bourgeon de membre, par
une petite tige de platine. Par suite de l'apparition tardive des corpuscules (elle
debute au 16-17eme jour d'incubation chez le Poulet et au 21eme jour chez le
Tableau 1. Types de greffes
realisees
et nombre de greffons etudies
histologiquement
Types de greffes
Greffes chorio-allantoidiennes
Greffes sur bourgeon frontal
Greffes dans le flanc
Greffes sur bourgeon de
membre
Greffes ccelomiques
Total
Nombre de greffons
etudies
73
24
11
216
113
437
Nombre de greffons
innerves
0
20
4
156
65
245
18-2

280R. SAXOD
frontal
Embryon donneur de tissu
nerveux, source d'innervation
Ganglion
spinal
Fig. 1. Greffe ccelomique par voie extraembryonnaire. Schemas de la face de
l'embryon donneur (Canard de 8 jours) et de la coupe transversale de l'embryon
note de
35
paires de somites au niveau des somites
20
a
25.
Le demi-bourgeon frontal
associe a une source d'innervation
(ici
un ganglion spinal) est introduit par une fente
dans
le coelome
extraembryonnaire puis pousse
en
direction mediane dans
le
coelome
embryonnaire.
Canard), les greffons ont ete preleves et fixes au liquide de Bouin vers la fin du
developpement embryonnaire du porte-greffe et parfois apres son eclosion.
Cette fixation tardive explique le faible pourcentage de cas etudies par rapport
au nombre de greffes realisees. Tous les greffons ont ete etudies histologique-
ment. Dans la serie des coupes, de 5 a
7
[im d'epaisseur, nous avons fait alterner
la coloration trichromique de Masson et la methode neurofibrillaire a l'uree-
nitrate d'argent (Ungewiter, 1951).

Le developpement des corpuscules de Herbst et de Grandry 281
Quatre types d'experiences ont ete realises (Tableau 1):
(1) Greffes chorio-allantoiidiennes: Poulet/Poulet, Canard/Poulet et Canard/
Canard (Tableau 2).
(2) Greffes orthotopiques (sur le bourgeon frontal, apres excision d'un frag-
ment d'ectoderme): Canard/Canard, Poulet/Canard et Canard/Poulet (Tableau
2) (Fig. 3).
(3) Greffes heterotopiques, dans le flanc ou sur le bourgeon de membre (apres
excision de sa moitie distale): Poulet/Poulet, Poulet/Canard, Canard/Poulet et
Canard/Canard (Tableaux 2, 3; Fig. 2). Des homogreffes de mesenchyme sous-
ectodermique1 de bourgeon frontal de Canard sur le bourgeon de membre de
Canard ont aussi ete realisees (en le glissant sous l'ectoderme de l'hote, ou en
l'appliquant contre la surface de section).
(4) Greffes coelomiques (Tableau 4; Figs. 1,4). Developpement de bourgeons
frontaux de Canard et de Poulet associes a diverses sources isolees d'innervation:
tube neural (moitie ventrale), ganglion sympathique (region sacree), ganglion
somato-sensitif (spinal ou cephalique d'embryons de Canard, de Poulet et de
Souris).
Ces greffes coelomiques sont realisees par voie extraembryonnaire. La tech-
nique est la suivante: sur des embryons ayant 30 a 35 paires de somites, une
incision de l'ectoderme et du mesoderme somatique est realisee au niveau des
arteres vitellines (22eme somite environ) a une distance de l'embryon egale a
environ quatre fois la largeur d'un somite. Le greffon est introduit par cette fente
dans le ccelome extraembryonnaire, puis pousse en direction mediane dans le
coelome embryonnaire. Dans la grande majorite des cas, les greffons sont
retrouves fixes sur le mesentere intestinal de l'embryon note et sont done exempts
d'innervation somatique. De plus, par rapport a la methode classique de greffe
ccelomique par incision dorsale de l'embryon, cette technique evite les hemor-
ragies et les lesions de l'embryon, elle ne provoque pas de coelosomie (plusieurs
eclosions ont ete obtenues) et rend possible l'introduction de greffons de grandes
dimensions.
RESULTATS
La differentiation morphologique et histologique de tous les greffons etudies
est conforme a leur origine (presence d'une ramphoteque et d'un axe ossifie
(Figs.
7, 8). Lorsqu'un demi-bourgeon frontral droit ou gauche est greffe, il se
produit toujours une regulation aboutissant a la formation d'un bee reduit mais
complet avec, chez le Canard, presence bilaterale de lamelles filtrantes (Figs.
2,4).
1 Le mesenchyme est separe de l'ectoderme a l'aide d'une solution de trypsine lyophilisee
a 0,5 % (Laboratoires Choay-France) et de pangestine a
1 %
(Difco) dans du liquide de Earle
sans calcium ni magnesium. Cette methode permet d'obtenir un mesenchyme exempt de
toute contamination ectodermique.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%