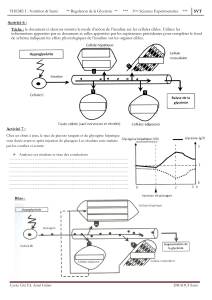
Régulation du métabolisme énergétique : hormones et hypoglycémie
Telechargé par
Svt pratique

Chapitre 18
VERSOPN IMPRIMABLE
REGULATION DU METABOLISME ENERGETIQUE
OBJECTIFS
De l’enseignant
Donner une vue plus précise de la régulation se référant à la régulation énergétique
Bien préciser les acteurs en jeu, hormones, réserves énergétiques, échanges tissulaires, etc.
Bien définir les hormones : structure et synthèse, mécanisme d’action, rôle métabolique, interactions et
antagonismes.
Montrer comment l’organisme fait face au problème vital de l’hypoglycémie grâce aux systèmes mis en œuvre.
Insister sur les dangers de l’hypoglycémie associée à l’alcoolisme.
De l’étudiant
Pour atteindre les objectifs de ce chapitre l’étudiant doit avoir assimilé les bases du métabolisme énergétique et
les interactions entre les différents tissus par le biais de leurs échanges de substrats. Ensuite ses objectifs seront les
suivants :
Bien connaître les acteurs en jeu, hormones, réserves énergétiques, échanges tissulaires, etc.
Bien Connaître les mécanismes d’action, rôle métabolique, interactions et antagonismes des hormones
impliquées.
Bien assimiler comment l’organisme fait face au problème vital de l’hypoglycémie grâce aux systèmes mis en
œuvre.
Prendre conscience des dangers de l’hypoglycémie associée à l’alcoolisme.
PLAN
OBJECTIFS
1 – INTRODUCTION
2 – HORMONES IMPLIQUEES DANS LA REGULATION DU METABOLISME ENERGETIQUE
2.1 - L’INSULINE
2.1.1 – STRUCTURE ET SYNTHESE
2.1.2 – SECRETION ET REGULATION
2.1.3 – MECANISME D’ACTION
2.1.4 – EFFETS METABOLIQUES
2.2 - LE GLUCAGON
2.2.1 - STRUCTURE
2.2.2 – STIMULATION ET INHIBITION DE LA SECRETION DU GLUCAGON
2.2.3 – MECANISME D’ACTION
2.2.4 – EFFETS METABOLIQUES
2.3 - L’ADRÉNALINE ET LA NORADRENALINE
2.3.1 - MECANISME D’ACTION
2.3.2 – EFFECTS METABOLIQUES
2.4 – CORTISOL
3 – HYPOGLYCEMIE ET SA GESTION MAR L’ORGANISME
3.1 - TYPES ET SYMPTOMES

3.1.1 - HYPOGLYCÉMIE POST PRANDIALE
3.1.2 - HYPOGLYCÉMIE DU JEÛNE
3.2 - LES SYSTEMES DE RÉGULATION DE LA GLYCEMIE
3.2.1 - SYSTÈME GLUCAGON-ADRÉNALINE
3.2.2 - SYSTÈME CORTISOL ET SUBSTANCES DE CROISSANCE
3.3 – HYPOGLYCEMIE ET L’ALCOOLISME
1 – INTRODUCTION
Le métabolisme énergétique des animaux se résume essentiellement de la façon suivante :
- Production de l’ATP et des précurseurs de biosynthèse à partir des glucides, des lipides et accessoirement des
protéines.
- Mise en réserve de l’énergie sous forme de glycogène et de triacylglycérols (triglycérides) lorsque
l’approvisionnement en aliments dépasse les besoins de l’organisme.
Ce métabolisme est soumis à une régulation complexe. Celle-ci met en jeu des hormones et prend en compte,
chez l’homme et les animaux, les réserves énergétiques, les échanges entre les tissus et les besoins journaliers de
l’organisme.La régulation est mise en œuvre lorsque se produit une inadéquation entre l’apport alimentaire journalier et
les besoins quotidiens de l’organisme.
Les réserves énergétiques chez un adulte de 70 kg se répartissent de la façon suivante :
- Glucides :
Glycogène hépatique : 100 g (6 à 8% du poids frais du foie)
Glycogène musculaire : 400 g (1 à 2% de la masse fraîche totale)
Glycogène sanguin : 20 g
- Lipides de réserve (triacylglycérols du tissu adipeux) : 13 000 à 14 000 g
- Protéines musculaires : 6000 g
Un régime est considéré comme équilibré si l’apport énergétique journalier contient :
- 60 % de glucides
- 30 % de lipides
- 10 % de protéines.
La régulation énergétique est aussi à l’origine des échanges entre les tissus suvant leurs états
nutritionnels pendant les périodes absorptives, post absorptives et le jeûne. Ces échanges ont fait l’objet du chapitre 17.
Dans ce nouveau chapitre nous allons nous intéresser aux hormones qui sont à la base de cette régulation énergétique,
à leur mécanisme et à leurs effets métaboliques.
OBJECTIFS 1 – INTRODUCTION 2 – HORMONES IMPLIQUEES
2.1 - L’INSULINE 2.2 - LE GLUCAGON 2.3 - L’ADRÉNALINE
2.4 – CORTISOL 3 – HYPOGLYCEMIE
2 – HORMONES IMPLIQUEES DANS LA REGULATION DU METABOLISME ENERGETIQUE
L’un des exemples les plus intéressants de régulation chez les animaux concerne la régulation du métabolisme
énergétique. La régulation de la glycémie, qui ajuste à chaque instant, le taux du glucose sanguin, est sous la
dépendance de trois hormones principales : l’insuline, le glucagon et l’adrénaline. L’Insuline est l’hormone majeure du
contrôle du métabolisme énergétique. Le glucagon , l’adrénaline et d’autres hormones comme la noradrénaline et le
cortisol ont des effets métaboliques destinés à contrecarrer ceux de l’insuline.
2.1 - L’INSULINE
L’Insuline est l’hormone majeure du contrôle du métabolisme énergétique. Elle coordonne l’utilisation des substrats
énergétiques. Ses effets sont anaboliques, favorisant les synthèses à savoir la glycogénogenèse, la lipogenèse et la
protéogenèse.
2.1.1 – STRUCTURE ET SYNTHESE

L’insuline est formée de 51 acides aminés, arrangés en deux chaînes polypeptidiques A (a) et B (b) reliées par 2
ponts disulfures. A l’intérieur de la chaîne A il existe un pont disulfure supplémentaire qui lie les résidus des acides
aminés 6 et 11. La structure de l’insuline humaine ne diffère de celle de l’insuline du porc que par l’acide aminé C
terminal de la chaîne B. Par hémisynthèse il suffit de remplacer l’alanine (C terminal) de la chaîne B de l’insuline du porc
par la thréonine pour obtenir l’insuline humaine.
La biosynthèse de l’insuline implique deux étapes intermédiaires conduisant à la formation de deux précurseurs
inactifs : préproinsuline et proinsuline (figure 1). Des clivages enzymatiques séquentiels fournissent l’insuline active, qui,
une fois synthétisée, est stockée dans des vésicules cytosoliques en attendant leur sécrétion par exocytose sous l’action
de stimuli spécifiques. La demi-vie de l’insuline dans le plasma est faible (environ 6 min), ce qui permet un ajustement
rapide de l’insuline dans le sang.
Figure 1 : Etapes de la synthèse de l’insuline et structure finale. Elle est synthétisée sous forme de préproinsuline qui
subit des clivages séquentiels qui l’amènent à sa structure définitive.
2.1.2 – SECRETION ET REGULATION
La production de l’insuline par les cellules b des îlots de Langerhans du pancréas est étroitement coordonnée à
celle du glucagon par les cellules a. La régulation du taux relatif de l’insuline et du glucagon contrôle l’aptitude du foie à
maintenir, à un niveau constant, l’approvisionnement en glucose des tissus périphériques. La production de l’insuline par
les cellules b répond à une variété de stimuli à savoir le glucose, les acides aminés, les hormones gastro-intestinales et
le glucagon.
Le glucose est le stimulus le plus important pour la production de l’insuline et auquel les cellules b sont les plus
sensibles chez les animaux. Un repas riche en glucides élève la sécrétion du l’insuline et diminue celle du
glucagon.
Les acides aminés, résultant de l’hydrolyse des protéines d’un repas, entraînent une sécrétion accrue de l’insuline.
L’acide aminé qui se comporte comme le stimulus le plus efficace est l’arginine.
Les hormones gastro-intestinales contribuent aussi à l’augmentation de la sécrétion de l’insuline à la suite d’un
repas. Elles ont un rôle d’anticipation, déclenchant la sécrétion de l’insuline avant l’effet de l’élévation sanguine en
glucose. Ceci permet d’expliquer pourquoi on observe une sécrétion d’insuline plus importante lorsqu’une même
quantité de glucose est administrée par voie orale que par voie veineuse.
Le glucagon voit sa sécrétion inhibée par le glucose pendant que ce dernier stimule celle de l’insuline. On en
conclut que la sécrétion de l’insuline inhibe celle du glucagon, ce qui constitue un contrôle du glucagon par
l’insuline. Dans le cas du diabète du type 1, plusieurs symptômes de la maladie seraient le fait des effets
métaboliques incontrôlés du glucagon, suite à la non sécrétion de l’insuline.
La synthèse et la sécrétion de l’insuline sont inhibées en l’absence de substrats énergétiques et en cas de
traumatisme. Dans ce dernier cas, le contrôle du métabolisme énergétique est récupéré par l’adrénaline sécrétée par la
médullosurrénale, en réponse au stress ou au traumatisme.
2.1.3 – MECANISME D’ACTION
L’insuline, une fois libérée, va se lier aux membranes des cellules-cibles qui possèdent des récepteurs
spécifiques transmembranaires du type catalytique. Chaque récepteur a deux domaines (figure 2) :
- un premier domaine extracellulaire qui fixe l’insuline (ligand) ;
- un second domaine intracellulaire, lié à l’activité tyrosine kinase.
La fixation de l'insuline, à son récepteur, active l’activité tyrosine kinase intrinsèque, qui phosphoryle des
tyrosines spécifiques au niveau des sous-unités b du récepteur (autophosphorylation) avant de phosphoryler les résidus

tyrosine des protéines cibles. Des phosphorylations en cascade sont initiées au niveau intracellulaire portant sur les
groupes hydroxyles des sérines des enzymes ou protéines cibles.
L’un des effets intracellulaires de l’insuline est l’activation par phosphorylation d’une protéine phosphatase.
Cette dernière, une fois activée, active ou inhibe, à son tour, par déphosphorylation, certaines enzymes-clés ou
protéines phosphorylées par la protéine kinase A, annihilant ainsi les effets antérieurs du glucagon ou de l’adrénaline.
On comprend ainsi les effets antagonistes entre l’insuline d’une part et les autres hormones d’autre part, intervenant
dant la régulation du métabolisme énergétique.
2.1.4 – EFFETS METABOLIQUES
Un repas riche en glucides augmente la teneur du sang en glucose en provenance de l’estomac et des intestins.
Ce signal est perçu et provoque une sécrétion de l’insuline en même temps qu’un ralentissement de la sécrétion du
glucagon. Ses effets sont très importants sur trois tissus : le foie, le muscle squelettique et le tissu adipeux. L’insuline est
responsable des effets métaboliques cités ci-après.
Métabolisme glucidique
· Dans le foie l’insuline inhibe la néoglucogenèse et la mobilisation du glycogène par l’inactivation de la
glycogène phosphorylase.
· Dans le muscle et dans le foie l’insuline active la synthèse du glycogène.
· Dans le muscle et le tissu adipeux l’insuline stimule l’entrée du glucose et accroît le nombre de transporteurs
transmembranaires du glucose (GLUT4). Le prélèvement accéléré du glucose dans le sang diminue la
glycémie et entraîne par voie de conséquence la baisse de la sécrétion de l’insuline

Figure 2 : Mécanisme d’action de l’insuline : La formation du complexe récepteur-insuline active une protéine kinase
tyrosine-spécifique qui assure une autophosphorylation du récepteur avant de phosphoryler le premier substrat cible
intracellulaire. Des réaction de phosphorylation en cascade conduisent à des effets intracellulaires.
Métabolisme lipidique
· Au niveau du tissu adipeux l’insuline inhibe la mobilisation des triglycérides en désactivant les enzymes
hormono-sensibles comme la triglycéride lipase, d’où la réduction du taux des acides gras dans le sang.
· L’insuline stimule la conversion de l’excès du glucose en acides gras. Elle accroît la synthèse des
triacylglycérols en activant les lipoprotéines lipases qui libèrent les acides gras. En favorisant l’entrée du
glucose et en stimulant la glycolyse dans les adipocytes, elle permet la fourniture du glycérol 3-, nécessaire
à la synthèse des triacylglycérols.
Métabolisme des acides aminés et des protéines
L’insuline favorise l’entrée des acides aminés en vue de la synthèse protéique dans la majorité des tissus
Par ses effets anaboliques l’insuline permet la reconstitution des réserves du glycogène dans le foie et dans les
muscles squelettiques, et la conversion des excès du glucose et des acides gras en triacylglycérols en vue de leur
stockage dans le tissu adipeux.
OBJECTIFS 1 – INTRODUCTION 2 – HORMONES IMPLIQUEES
2.1 - L’INSULINE 2.2 - LE GLUCAGON 2.3 - L’ADRÉNALINE
2.4 – CORTISOL 3 – HYPOGLYCEMIE
2.2 - LE GLUCAGON
L’utilisation continue du glucose par le cerveau et les autres tissus entraîne, au bout de quelques heures après un
repas même en l’absence d’effort physique, un abaissement du taux du glucose au-dessous de 4.5 mM. La chute du
glucose à ce niveau déclenche la synthèse et la sécrétion du glucagon et ralentit celle de l’insuline.
2.2.1 - STRUCTURE
Le glucagon est une hormone peptidique, synthétisée par les cellules a des îlots de Langerhans du pancréas. Il
est constitué de 29 acides aminés, arrangés dans une seule chaîne peptidique. Contrairement à l’insuline, la séquence
peptidique est la même chez tous les mammifères, dont les séquences ont été analysées à ce jour. Il est synthétisé sous
forme d’un précurseur, qui, par clivages séquentiels, fournit l’hormone active.
2.2.2 – STIMULATION ET INHIBITION DE LA SECRETION DU GLUCAGON
Les cellules a répondent à des signaux indiquant une hypoglycémie ou destinés à anticiper des besoins en
glucose par la sécrétion du glucagon. Les stimuli sont variés : abaissement du taux du glucose sanguin, acides aminés,
adréaline, etc.
· L’abaissement du taux de glucose plasmatique est le premier stimulus pour la sécrétion du glucagon. Pendant la
phase post-absorptive et pendant le jeûne prolongé c’est le glucagon qui permet de maintenir la glycémie à un
niveau constant.
· Après un repas riche en protéines, les acides aminés issus de leur hydrolyse stimulent à la fois la sécrétion du
glucagon et celle de l’insuline. Le glucagon contrecarre les effets de l’insuline, qui entraîneraient une
hypoglycémie par prélèvement du glucose,
· L’adrénaline, provenant de la médullo-surrénale, et la noradrénaline, produite par le système sympathique
innervant le pancréas, activent la sécrétion du glucagon. Grâce à l’action de ces deux dernières hormones, le
taux élevé du glucagon dans le sang permet à l’organisme de répondre aux besoins en glucose pendant le
stress et les traumatismes ou d’anticiper ses besoins en cas d’efforts physiques mportants.
La sécrétion du glucagon est inhibée par une forte concentration du glucose dans le sang et par l’insuline après
un repas riche en glucides.
 6
7
8
9
10
6
7
8
9
10
1
/
10
100%