Cours de Biologie Végétale : Algues, Bryophytes, Ptéridophytes
Telechargé par
lindaylasme31

République de Côte d’Ivoire_Union-Discipline-Travail
Minsitère de l’Enseignement Supérieur et de la Recherche Scientifique
Classes Préparatoires Biologiques 2ème année
Support de cours
Support de coursSupport de cours
Support de cours
BIOLOGIE VEGETALE
BIOLOGIE VEGETALEBIOLOGIE VEGETALE
BIOLOGIE VEGETALE
Proposé par :
INSTITUT NATIONAL POLYTECHNIQUE FELIX HOUPHOUËT-BOIGNY
ECOLE PREPARATOIRE
Année académique : 2010-2011
KOUAKOU Kouadio Paul
KOUAKOU Kouadio PaulKOUAKOU Kouadio Paul
KOUAKOU Kouadio Paul
-
--
-
Martial
MartialMartial
Martial
Ingénieur Agronome (32ème Promotion ENSA)
Enseignant-Chercheur / Assistant
Laboratoire de Recherche en Sciences Agronomiques et Génie Rural
Département Agriculture et Ressources Animales
© Septembre 2010
Prof BROU Yao Casimir
Prof BROU Yao CasimirProf BROU Yao Casimir
Prof BROU Yao Casimir
Maître de Conférence
Enseignant-Chercheur
Laboratoire de Recherche en Sciences Agronomiques et Génie Rural
Département Agr
iculture et Ressources Animales

SOMMAIRE
SOMMAIRESOMMAIRE
SOMMAIRE
Page
INTRODUCTION
INTRODUCTIONINTRODUCTION
INTRODUCTION
1
11
1
I. ORGANISMES VIVANTS A STRUCTURE CELLULAIRE…………………….………………. 1
II. DIFFERENCES ENTRE CORMOPHYTES ET THALLOPHYTES…………………………..…... 1
III. CLASSIFICATION DES VEGETAUX……………………………………….………………. 2
IV. ELEMENTS DE PHYLOGENIE……………………………………………………………... 6
Chapitre I
Chapitre IChapitre I
Chapitre I
: REGNE DES PROTISTES
: REGNE DES PROTISTES: REGNE DES PROTISTES
: REGNE DES PROTISTES-
--
- EMBRANCHEMENT DES ALGUES
EMBRANCHEMENT DES ALGUES EMBRANCHEMENT DES ALGUES
EMBRANCHEMENT DES ALGUES
7
77
7
A. ETUDE DES DIFFERENTS EMBRANCHEMENTS……………………………
A. ETUDE DES DIFFERENTS EMBRANCHEMENTS……………………………A. ETUDE DES DIFFERENTS EMBRANCHEMENTS……………………………
A. ETUDE DES DIFFERENTS EMBRANCHEMENTS………………………………..
…..…..
…..…………..
…………..…………..
…………..
7
77
7
INTRODUCTION………………………………………………………………….……………
7
I. CLASSE DES PYRRHOPHYTES………………………………………………………………..
8
II. CLASSE DES CHRYSOPHYTES……………………………………..........................................
8
III. CLASSE DES BACILLARIOPHYTES………………………………………………………….
9
IV. CLASSE DES EUGLENOPHYTES…………………………………………………………….
10
V. CLASSE DES CHLOROPHYTES……………………………………………………………... 11
VI. CLASSE DES PHEOPHYTES…………………………………………………………………
13
VII. CLASSE DES RHODOPHYTES……………………………………........................................
13
VIII. INTERET DES ALGUES…………………………………………………………………… 14
B. REPRODUCTION ET CYCLE DE DEVELOPPEMENT DES ALGUES………
B. REPRODUCTION ET CYCLE DE DEVELOPPEMENT DES ALGUES………B. REPRODUCTION ET CYCLE DE DEVELOPPEMENT DES ALGUES………
B. REPRODUCTION ET CYCLE DE DEVELOPPEMENT DES ALGUES………………
………………
…………
……
………….
……….……….
……….
1
11
15
55
5
I. GENERALITES………………………………………………………………………………...
15
1.1. Reproduction asexuée……………………………………………………...……… 15
1.2. Reproduction sexuée………………………………………………………………. 15
1.3. Cycle de développement……………………………………………………………
16
II. ETUDE D’UNE CHLOROPHYTE FILAMENTEUSE : LA SPYROGYRE……………...................
17
III. ETUDE D’UNE CHLOROPHYTE NON FILAMENTEUSE : L’ULVE…………………………. 18
3.1. Présentation………………………………………………………………………... 18
3.2. Reproduction et cycle de développement…………………………………………. 18
IV. ETUDE D’UNE PHEOPHYTE : UNE LAMINAIRE………………..………………………… 19
V. CYCLE DE DEVELOPPEMENT D’UNE RHODOPHYCEE :
ANTITHAMNION PLUMULA.....
..
20
Chapitre
Chapitre Chapitre
Chapitre I
II
II
II
I
:
: :
: REGNE DES VEGETAUX
REGNE DES VEGETAUXREGNE DES VEGETAUX
REGNE DES VEGETAUX-
--
-
EMBRANCHEMENT DES BRYOPHYTES
EMBRANCHEMENT DES BRYOPHYTESEMBRANCHEMENT DES BRYOPHYTES
EMBRANCHEMENT DES BRYOPHYTES
22
2222
22
I. GENERALITES………………………………………………………...………………………
22
1.1. Embranchement des Bryophytes………………………………………..………….
22
1.2. Cycle de développement ou cycle de vie…………………………………………...
22
1.3. Mousse acrocarpe- Mousse pleurocarpe…………………………………………..
24
II. CLASSIFICATION DES BRYOPHYTES……………………………………………………….
25
2.1. Classe des Mousses…………………………………………………………………
25
2.2. Classe des Hépatiques………………………………………………………………
27

2.3. Classe des Anthocérotes………………………………..…………………………...
28
III. INTERET DES BRYOPHYTES………………………………………………...……………...
28
Chapitre II
Chapitre IIChapitre II
Chapitre III
II
I
:
: :
: REGNE DES VEGETAUX
REGNE DES VEGETAUXREGNE DES VEGETAUX
REGNE DES VEGETAUX-
--
-
EMBRANCHEMENT DES PTERIDOPHYTES
EMBRANCHEMENT DES PTERIDOPHYTESEMBRANCHEMENT DES PTERIDOPHYTES
EMBRANCHEMENT DES PTERIDOPHYTES
29
2929
29
I. GENERALITES…………………………………………………………………...……………
29
II. ORGANISATION DU GAMETOPHYTE……………………………………………………...
30
2.1. Prothalle………………………………………………………………......................
30
2.2. Organes reproducteurs………………………………………………......................
31
III. ORGANISATION DU SPOROPHYTE…………………………………………...................... 31
3.1. Racine…………………………………………………………………..................... 31
3.2. Tige……………………………………………………………………..................... 32
3.3. Feuille ou fronde……………………………………………………………………
33
3.4. Sporanges et spores………………………………………………………………... 33
3.5. Sexualité des spores………………………………………………………………...
34
IV. CLASSIFICATION DES PTERIDOPHYES…………………………………………………… 35
4.1. Classe des Psilotinées…………………………………………………......................
36
4.2. Classe des Lycopodinées……………………………………………………………
37
4.3. Classe des Equisétinées (Articulées, Prêles)………………………………………...
38
4.4. Classe des Filicinées………………………………………………………………...
39
V. IMPORTANCE DES PTERIDOPHYTES……………………………………………………… 39
Chapitre I
Chapitre IChapitre I
Chapitre IV
VV
V
:
: :
: REGNE DES VEGETAUX
REGNE DES VEGETAUXREGNE DES VEGETAUX
REGNE DES VEGETAUX-
--
-
EMBRANCHEMENT DES SPERMAPHYTES
EMBRANCHEMENT DES SPERMAPHYTESEMBRANCHEMENT DES SPERMAPHYTES
EMBRANCHEMENT DES SPERMAPHYTES
40
4040
40
INTRODUCTION……………………………………………………………………………… 40
A
AA
A-
--
-
SOUS
SOUSSOUS
SOUS-
--
-EMBRANCHEMENT DES GYMNOSPERMES………………………………...
EMBRANCHEMENT DES GYMNOSPERMES………………………………...EMBRANCHEMENT DES GYMNOSPERMES………………………………...
EMBRANCHEMENT DES GYMNOSPERMES………………………………...............
........................
............
41
4141
41
I. GENERALITES………………………………………………………………………………...
41
II. CLASSIFICATION ET CARACTERES ESSENTIELS DES GYMNOSPERMES………………….. 41
2.1. Classe des Cycadophytes………………………………………………....................
42
2.2. Classe des Coniférophytes………………………………………………………….
42
2.3. Classe des Gnétophytes……………………………………………………………..
43
III. CYCLE DE DEVELOPPEMENT……………………………………………………………... 44
3.1. Etude d’une Gymnosperme primitive (le Cycas)…………………………………..
44
3.2. Etude d’une Gymnosperme type (le pin sylvestre)……………………....................
45
B
BB
B-
--
- SOUS
SOUS SOUS
SOUS-
--
-EMBRANCHEMENT DES ANGIOSPERMES………………………………
EMBRANCHEMENT DES ANGIOSPERMES………………………………EMBRANCHEMENT DES ANGIOSPERMES………………………………
EMBRANCHEMENT DES ANGIOSPERMES………………………………...
......
....
..
...............
............................
..............
4
44
48
88
8
I. GENERALITES………………………………………………………………………………...
48
II. FLEUR DES ANGIOSPERMES……………………………………………………………….. 49
III. FRUIT DES ANGIOSPERMES……………………………………………………………......
51
IV. CYCLE DE DEVELOPPEMENT DES ANGIOSPERMES………………………………………
51

Introduction
1
INTRODUCTION
I. ORGANISMES VIVANTS A STRUCTURE CELLULAIRE
Selon la classification adoptée par Whittaker (1969), les organismes vivants à structure cellulaire ont été
subdivisés en 5 grands règnes:
Le règne des Monères (Monera, Procaryotes) ;
Le règne des Protistes (Protista) ;
Le règne des Mycètes (Fungi, Champignons) ;
Le règne des Végétaux (Plantae, Plantes) ;
Et le règne des Animaux (Animalia).
(Voir Tableau).
Spécificité des végétaux
Les végétaux sont tous eucaryotes donc compartimentés (organites). Chaque organite joue un rôle dans
la cellule. Il y a spécialisation des organites pour une certaine fonction, cette spécialisation permet une
meilleure efficacité ;
Ils sont pluricellulaires ;
Ils sont libres ou fixés ;
Ils sont pour la plupart autotrophes mais peuvent être aussi hétérotrophes (quelques espèces) ;
Les cellules végétales ont une paroi qui se situe à l'extérieur de la membrane et faite de polysaccharides
et, est en général rigide, permettant le maintien du végétal (qui est sans squelette). Cette paroi possède
des petits orifices : les plasmodesmes qui permettent la communication et les échanges intercellulaires ;
Les plastes sont des organites spécifiques des végétaux (chloroplastes, amyloplastes (amidon),
chromoplastes (pigments)) ;
Chez les végétaux, on trouve à l'intérieur de la cellule une vacuole. C'est une poche entourée d'une
membrane pleine d'eau et de formes ioniques. Elle permet elle aussi le port du végétal.
II. DIFFERENCES ENTRE CORMOPHYTES ET THALLOPHYTES
Les végétaux étaient anciennement subdivisés en Cormophytes (végétaux supérieurs) et en Thallophytes
(végétaux inférieurs). Les Cormophytes forment un grand groupe qui se distingue par sa morphologie
(forme, aspect extérieur), sa structure anatomique (organisation interne : différenciation cellulaire et
agencement de tissus) et par la structure de ses organes reproducteurs, par rapport aux Thallophytes.

Introduction
2
2.1. Au plan morphologique
Sur le plan morphologique, les Cormophytes se caractérisent par un appareil végétatif qui dans le cas typique
est constitué d'un cormus (tige ou tronc) portant latéralement des feuilles (organes aériens aplatis) et des
racines (organes souterrains). Les Bryophytes sont les seuls végétaux dépourvus de racines et qui possèdent
seulement des rhizoïdes.
Les Cormophytes sont généralement ou essentiellement des végétaux terrestres par opposition aux
Thallophytes qui sont généralement aquatiques.
Les Cormophytes sont des organismes sporophytiques (organismes diploïdes à l'exception des Bryophytes
qui sont des organismes gamétophytiques et donc des organismes haploïdes).
Les Cormophytes sont des végétaux chlorophylliens (à l’exception de certaines plantes parasites) et sont de
ce fait autotrophes avec une croissance indéfinie (grâce aux méristèmes apicaux) ; ce qui les opposent
fondamentalement aux organismes du monde animal qui sont hétérotrophes et à croissance définie tels que
les vertébrés.
2.2. Au plan anatomique
Sur le plan anatomique, les Cormophytes possèdent (à l'exception des Bryophytes) une structure différenciée
avec des tissus spécialisés remplissant différents rôles et fonctions : tissus protecteurs (épiderme), tissus
assimilateurs (pour la photosynthèse), tissus conducteurs (conduction de la sève), etc.
2.3. Au plan de la reproduction
Sur le plan de la reproduction, les Cormophytes possèdent des organes reproducteurs muni d’une enveloppe
pluricellulaire qui entoure les cellules reproductrices (gamètes, spores) et de ce fait on parle de gamétanges
pour les organes contenant les gamètes et de sporanges pour les organes contenant les spores, contrairement
aux organes des Thallophytes (gamétocystes et de sporocystes) qui ne sont entourés que par la paroi de la
cellule mère.
Une autre caractéristique des Cormophytes réside dans la structure de leur gamétange femelle ou archégone :
organe en forme de bouteille constitué d'un ventre, un col et un canal. A cause de cette structure typique
(archégone), le terme Archégoniates est utilisé le plus souvent pour désigner les Cormophytes (Cormophytes
= Archégoniates).
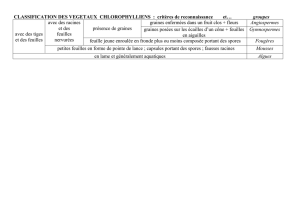
III. CLASSIFICATION DES VEGETAUX
Cette classification est simplifiée et vise à donner les grands groupes de Cormophytes et à les situer les uns
par rapport aux autres. Certaines caractéristiques sont aussi précisées.
3.1. Critères taxonomiques
Le groupe des Cormophytes comprend (sommairement) 4 grands Embranchements. Des moins évolués aux
plus évolués on distingue :
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
1
/
57
100%