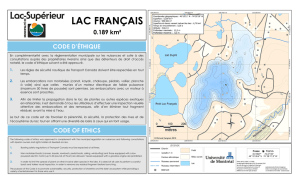
role des facteurs écologiques dans l`évolution

LYSANDRE LANDRY
ROLE DES FACTEURS ÉCOLOGIQUES DANS
L'ÉVOLUTION PARALLÈLE DU GRAND COREGONE
(COREGONUS CLUPEAFORMIS MiTCHILL.)
Mémoire présenté
à la Faculté des études supérieures de l'Université Laval
dans le cadre du programme de maîtrise en biologie
pour l'obtention du grade de Maître es Sciences (M. Se.)
DÉPARTEMENT DE BIOLOGIE
FACULTÉ DES SCIENCES ET DE GÉNIE
UNIVERSITÉ LAVAL
QUÉBEC
JUIN 2006
Lysandre Landry, 2006

RÉSUMÉ
La sélection naturelle est un des facteurs dominants lors de l'évolution répétée et parallèle
observée notamment chez les poissons des régions tempérées du Nord. Ainsi, au sein des
populations de Coregonus clupeaformis, l'écotype nain
s'est
différencié plusieurs fois à
partir d'un écotype normal, notamment dans six lacs du bassin versant de la rivière St-Jean
(Maine, USA et Québec, CA). Cette étude porte sur les caractéristiques de l'habitat ainsi
que sur la disponibilité des ressources et leurs variations temporelles influençant le niveau
de différenciation atteint entre les écotypes de ces populations. Les résultats appuient les
hypothèses théoriques faisant intervenir le rôle des fluctuations environnementales
(biomasse et structure des ressources) dans la variation des pressions sélectives et le rôle de
la compétition pour les ressources (disponibilités réduites) dans l'établissement et le
maintien d'un degré de différenciation. Le rôle de d'autres interactions trophiques comme
les relations prédateurs-proies semble aussi impliqué dans ce mécanisme de divergence.
ABSTRACT
Natural sélection is one of the dominant factors in repeated parallel évolution observed
notably in north temperate fish populations. In Coregonus clupeaformis populations located
in six lakes of the St-John's.river watershed (Maine, USA and Québec, CA), a dwarf
ecotype has differentiated from a normal ecotype many times. Hère we study habitat and
resource features, it temporal variations in relation to the observed level of differentiation
between écotypes of thèse populations. The results support the hypothèses that
environmental fluctuation (zooplankton community structure and biomass) cause variation
in sélective pressure and that the resource compétition (availability of habitat and
resources) influence the establishment and persistence of a degree of differentiation.
Trophic interactions such as predator-prey relationships seem also to be involved in giving
rise to this divergence.

11
AVANT-PROPOS
De l'exaltation de mes premiers pas dans le laboratoire du docteur Louis Bernatchez,
jusqu'à la fin de ce très long et cahoteux chemin, j'ai rencontré une multitude de personnes
merveilleuses et généreuses avec qui j'ai grandi et sans qui je n'y serais jamais arrivée. Je
veux d'abord remercier Louis Bernatchez pour sa patience et sa confiance en moi ainsi que
Sean M. Rogers et David Campbell; les premiers qui m'ont fait vibrer par leur passion pour
la biologie évolutive. Ensuite et non moindrement, merci aux personnes qui ont fait et qui
font encore partie de l'équipe du projet corégone, sans oublier Vicky Albert sans qui je
n'aurais jamais fait partie de ce projet. Vive la plus belle «gang» évolutive au monde!!!
Je veux remercier aussi à mon co-directeur Warwick F. Vincent pour son soutien et ses
encouragements ainsi que les membres de son laboratoire : Sébastien Roy, Leira Rétamai,
Marie-Josée Martineau et Milla Rautio pour l'aide en limnologie.
Toute ma gratitude à Gesche Winkler qui m'a soutenue, rassurée et aidée dans le choix des
meilleurs protocoles pour traiter les données d'invertébrés lacustres, pour les discussions
écologiques et plus encore. Merci à Julian J. Dodson et aux gens de son laboratoire qui ont
toujours été là quand j'en avais besoin.
Merci à Robert St-Laurent sans qui les gens qui m'ont aidé et accueilli sur le terrain
n'aurait jamais été aussi heureux, mais aussi pour son aide avec le matériel de terrain et
avec l'échantillonnage, et pour les discussions passionnées qui m'ont permis de continuer.
Merci à la contribution généreuse des biologistes du «Maine Department of Inland
Fisheries and Wildlife»; M. David Basley et ses heureux acolytes Derrick Cote et Frank O.
Frost, qui m'ont permis d'obtenir des invertébrés à observer et des échantillons à analyser.
Je veux remercier aussi les aubergistes du Lac de l'Est pour nous avoir accueilli et permis
d'utiliser leur espace de vie pour les observations et analyses sur le terrain.
Merci à mes joyeux compagnons de terrain, de tri et d'identification : Indi Noël, Robin
Martel, Julie Breton, Justin Roy, Guillaume Côté et Tanguy Castric qui ont survécu à mes
angoisses benthiques et bien sûr, à Lucie Papillon pour sa compréhension et son sens de
l'organisation.

111
Merci aux «cracs» des statistiques, des mathématiques et de la réflexion logique: Pierre
Duchesne, Julie Béliveau, Sophie Baillargeon, Scott McCairns, Mathieu Cusson, Milla
Rautio et David Véliz.
Je veux remercier particulièrement Guillaume Côté, Mélissa Lieutenant-Gosselin, Scott
McCairns, Julien April et Anne-Marie Gale ainsi que ma petite soeur Mylène et mon père
Jacques Landry pour leur patience et leur contribution généreuse à l'amélioration de ce
mémoire.
Finalement, je veux remercier encore Sylvain Gutjahr, Suzy Tremblay, Guillaume Côté,
Mélissa Lieutenant-Gosselin, Scott McCairns, Marie-France Barrette, Véronique Thériault,
Pierre-Philippe Dupont, Vincent Bourret, Vanessa Viera, Sean Rogers, David Campbell,
David Véliz, Catherine Lippe, Dylan J. Fraser, Mélanie Dionne, Dany Garant, Mathieu
Cusson, Catherine Potvin, Marc Ringuette et Frédéric Lecomte pour les discussions
scientifiques et personnelles passionnantes, pour leurs conseils et leur confiance en moi. Je
veux remercier ma mère Paule Croteau, mes sœurs Mylène et Marie-Christine, mon père
Jacques Landry et Kerstin Bellmann pour le soutien et l'affection.
L'article qui compose le cœur de ce mémoire a été soumit au journal Evolution. Le travail
de laboratoire et l'analyse des données ont été réalisés par L. Landry sous la supervision du
directeur, L. Bernatchez et du co-directeur, W. F. Vincent. Une partie du travail de
laboratoire a été fait avec l'aide de R. Martel. La rédaction a été effectuée avec l'aide et les
critiques de L. Bernatchez et de W. F. Vincent. De plus, j'ai également participé à une
étude sur la spécialisation alimentaire des écotypes nains et normaux du Grand Corégone à
l'aide d'isotopes stables. Le travail de laboratoire et l'échantillonnage ont été réalisés par
N.
Pelletier et l'analyse des données par L. Landry. La rédaction de ces travaux sera
effectuée par L. Landry sous la supervision du directeur L. Bernatchez. J'ai aussi effectué
des travaux sur la faune benthique dont le travail au laboratoire a été fait avec l'aide de
Julie Breton et de Indi Noël. Je ferai l'analyse de ces données et je participerai à la
rédaction d'une publication sur celles-ci avec l'aide de L. Bernatchez. Finalement, L.
Landry a été soutenu financièrement par le groupe de recherches inter-universitaires,
Québec-Océan.

«Les choses que je sens et que
j'imagine
ne sont
peut-être rien du tout hors de moi et en elles-
mêmes». Descartes, Deuxième méditation
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
1
/
68
100%