Sida, des bases pour comprendre - Act Up

information
=pouvoir
2
ACT UP PARISTMjanvier 2010
SIDA, DES BASES POUR COMPRENDRE 2010
information
= pouvoir
2SIDA, DES BASES POUR COMPRENDRE
Act Up-Paris est une association issue de la communauté homosexuelle,
veillant à défendre équitablement toutes les populations touchées par le sida.
Act Up-Paris n’est pas une association caritative, mais une association de
personnes concernées par le VIH qui, au delà de leur tragédie personnelle,
voient dans le sida avant tout une question politique.
Act Up-Paris porte sur la place publique un discours et des exigences de per-
sonnes atteintes. Ce discours est complémentaire et parfois contraire à celui
des médecins, des pouvoirs publics ou des laboratoires pharmaceutiques.
Act Up-Paris a aussi pour mission d’informer les malades sur les traitements
disponibles, afin qu’ils, elles, participent en connaissance de cause à la déci-
sion thérapeutique. Parce que l’information est un pouvoir.
Act Up-Paris a été créé en 1989 par Didier Lestrade, Pascal Loubet et Luc
Coulavin. Depuis 20 ans maintenant notre association a contribué à changer
radicalement le cours de l’épidémie de sida : en devenant un vrai outil de pres-
sion activiste, en faisant évoluer le discours sur la maladie, en révélant et com-
battant toute forme de discrimination envers les personnes touchées et leur
entourage. Depuis 1995, nous avons été au premier rang des associations de
lutte contre le sida pour que certains antirétroviraux soient délivrés à un grand
nombre de malades avant leur commercialisation effective. Résultat de ce har-
cèlement constant : la France est le pays européen où les nouvelles molécu-
les et les trithérapies sont le plus largement accessibles. Plus de 110 000 per-
sonnes suivent un traitement antirétroviral.
Mais tout n’est pas réglé. D’autres médicaments sont en cours de développe-
ment car des échecs thérapeutiques continuent à se produire : près de 8 %
des personnes traitées ne répondent pas aux trithérapies réputées les plus
efficaces. Nous devons continuer notre pression pour que les recherches de
nouveaux médicaments se poursuivent, mais dans le respect des règles éthi-
ques établies. Nous devons développer l’information sur ces nouveaux traite-
ments auprès des personnes atteintes. Nous devons nous battre pour que la
co-infection VIH/hépatite soit prise en compte par tous les acteurs de la lutte
contre le sida. Nous avons besoin de votre aide.
Téléphonez-nous pour connaître les horaires de nos groupes de travail ou venez à nos réunions
tous les jeudis soirs, 19h00, à l’Ecole des Beaux Arts, 14 rue Bonaparte, Paris VIe.
Pour nous contacter par mail, faites précéder [@actupparis.org] du nom lié au sujet de votre
envoi parmi la liste ci-après :
accueil • actup • actupinfos • administration • coinfection • communication • coordination-ds •
comptabilite • diffusion • drogues • droits • etrangers • events • femmes • financement • gap •
groupcomm • homophobie • international • jeunes • medias • permanence • plaidoyersud • pre-
vention • prison • prostitution • publications • reactup • revuedepresse • secretariat • sidablabla
• stands • traitements • web
Information=pouvoir est une collection unique qui rassemble et actualise
l’expertise des militantEs d’Act Up-Paris. Depuis maintenant 20 ans, Act Up-
Paris veille à défendre équitablement toutes les populations touchées par le
sida.
En France, chaque année, des milliers de personnes découvrent leur
séropositivité, malgré les messages de prévention largement diffusés.
Act Up-Paris porte sur la place publique une information sans concession,
qui permet la meilleure autonomie de décision du public concerné.
Le numéro 1(SIDA, UN GLOSSAIRE) est disponible à Act Up-Paris.
Les 2(DES BASES POUR COMPRENDRE) et 3(GUIDE DES DROITS SOCIAUX) sont
diffusés en librairie par les éditions La Découverte, qui assurent ainsi
un accès élargi à des ouvrages indispensables.
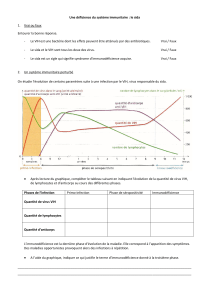
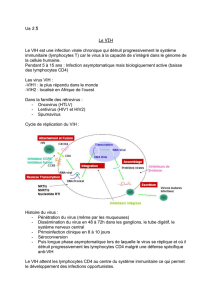
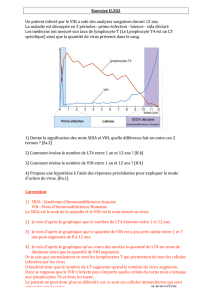
SIDA, DES BASES POUR COMPRENDRE donne une lisibilité exceptionnelle
aux phénomènes biologiques complexes en jeu dans la maladie.
En particulier, 58 illustrations inédites et novatrices permettent
de visualiser clairement l’essentiel.
Ce Guide est réalisé par les militantEs d’Act Up-
Paris. Il est tiré à 15 000 exemplaires. Il ne peut
être vendu. La reproduction des articles est
autorisée à condition d’en citer la source :
SIDA
,
DES BASES POUR COMPRENDRE
, d’Act Up-Paris

Depuis sa création, Act Up-Paris revendique l’accès à l’information.
Être informé n’est pas un devoir, mais la possibilité d’agir sur le cours des choses.
Pour permettre à chacunE de lutter efficacement contre le sida et de puiser aux meilleures sour-
ces l’information dont il/elle a besoin, nous éditons :
Action bulletin d’information sur les enjeux de la lutte contre le sida.
Abonnement : 5 numéros par an - 8 euros ou 18 euros (soutien).
Protocoles bulletin d’information thérapeutique sur l’actualité médicale et scientifique, les trai-
tements contre le VIH et contre les hépatites virales et les essais cliniques.
Abonnement : 5 numéros par an - 8 euros.
Pour toutes questions concernant les abonnements ou la diffusion des guides, contactez Rose
Rachel Rebelle : [email protected]g - 01 49 29 44 78
www.actupparis.org retrouvez toutes nos publications, les comptes-rendus de nos Réunions
Publiques d’Information (RéPI), nos communiqués et dossiers de presse, etc.
collection information = pouvoir
1SIDA, UN GLOSSAIRE
a pour but de mieux comprendre les termes médicaux, d’interpréter son bilan sanguin,
de participer plus activement au dialogue avec son médecin.
La cinquième édition est parue en décembre 2009.
2SIDA, DES BASES POUR COMPRENDRE
explique toutes les bases de la science aux personnes qui veulent comprendre la recherche
scientifique de la lutte contre le sida. La première édition est parue en mars 2010.
3 SIDA, LE GUIDE DES DROITS SOCIAUX
présente une information claire et précise de tous les droits auxquels peuvent prétendre les per-
sonnes séropositives. La troisième édition est parue en avril 2010.

information
=pouvoir
2
SIDA, DES BASES POUR COMPRENDRE
ACT UP-PARISTMJANVIER 2010

des bases pour comprendre
l’ensemble des illustrations et des graphiques
sont conçus et réalisés par Emmanuel Baume et Franck Poulet
conception maquette, suivi réalisation avec Claire Vannier : Carole Peclers
table des matières exhaustive p. 197 selon la mise en ligne
www.actupparis.org

biologie
le système immunitaire
l’infection à VIH
1
2
3
4
5
6
outils supplémentaires
prévention de la transmission du VIH
la recherche clinique
1.1 concepts
1.2 machinerie cellulaire
1.3 le génome
1.4 la transcription
1.5 la traduction
2.1 les organes du système immunitaire
2.2 origines et cellules souches
2.3 immunité innée
2.4 immunité acquise ou adaptative
2.5 entre inné et acquis
3.1 le virus de l’immunodéficience humaine
3.2 mécanisme de la maladie
3.3 les antirétroviraux
4.1 parcours du médicament
4.2 résisitances du virus
4.3 origine et histoire
5.1 dépistage et diagnostic
5.2 transmission sexuelle du
VIH
5.3 antirétroviraux pour la prévention
6.1 comprendre les essais cliniques
6.2 participer à un essai clinique
6.3 le consentement éclairé
6.4 quand un essai s’arrête
6.5 autres moyens d’accéder à de nouveaux traitements
6.6 quelles questions dois-je me poser ?
6.7 comment un essai clinique voit-il le jour ?
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
1
/
202
100%