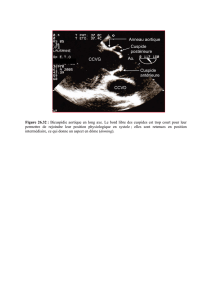
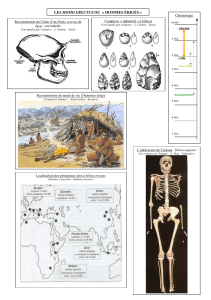
I LES DIFFERENTS THEORIES DE L`EVOLUTION

UNIVERSITE DE NANTES
UNITE DE FORMATION ET DE RECHERCHE D’ODONTOLOGIE
Année 2007 Thèse n°13
THESE
Pour le Diplôme D’Etat de
DOCTEUR en CHIRURGIE DENTAIRE
Par
Florie DURANTEAU
Née le 05 octobre 1979
Présentée et soutenue publiquement le 13 mars 2007
EVOLUTION MORPHOLOGIQUE DES DENTS
DES ANTHOPOIDES A L’HOMME
JURY
Président : Monsieur le Professeur A. JEAN
Assesseur : Monsieur le Professeur B. GIUMELLI
Assesseur : Monsieur le Professeur O. LABOUX
Directeur : Monsieur le Docteur A. HOORNAERT
1

INTRODUCTION _______________________________________________ 5
1 LES DIFFERENTES THEORIES DE L’EVOLUTION ______________ 6
1. L’évolution des espèces _______________________________________ 6
1. Le Fixisme (XIXème siècle) ___________________________________ 6
2. Le Transformisme (XIXème siècle) _____________________________ 7
3. L’Evolutionnisme (XIXème siècle) _____________________________ 8
4. La Récapitulation (XIXème siècle)______________________________ 8
5. Le Néo Darwinisme ou théorie synthétique de l’évolution (XXème siècle)
___________________________________________________________ 9
6. Les Equilibres Ponctués (XXème siècle) ________________________ 10
2. Evolution et origines de l’homme______________________________ 11
1. East side story ____________________________________________ 11
2. La biodynamique cranio-faciale_______________________________ 11
3. La bipédie originelle (21)____________________________________ 13
2 DE TOUMAI A HOMO SAPIENS : DESCRIPTION DE LA FACE DE LA
MANDIBULE ET DES DENTS___________________________________ 15
1. Toumaï (Sahelanthropus Tchadensis)__________________________ 16
2. Orrorin Tungenensis________________________________________ 17
3. Les Australopithèques_______________________________________ 19
1. Ardipithecus Ramidus ______________________________________ 19
2

2. Australopithecus Anamensis _________________________________ 20
3. Australopithecus Afarensis __________________________________ 24
4. Kenyanthropus Platyops ____________________________________ 29
5. Australopithecus Africanus __________________________________ 30
6. Australopithecus Bahrelghazali _______________________________ 33
7. Australopithecus Gahri _____________________________________ 34
4. Les Premiers Hommes ______________________________________ 36
1. Paranthropus Aethiopicus ___________________________________ 36
2. Homo Habilis_____________________________________________ 39
3. Homo Rudolfensis _________________________________________ 41
4. Paranthropus Boisei ________________________________________ 42
5. Paranthropus Robustus______________________________________ 47
6. Homo Ergaster ____________________________________________ 49
7. Homo Erectus_____________________________________________ 51
8. Homo Heidelbergensis______________________________________ 54
9. Homo Néanderthalensis_____________________________________ 61
10. Homo Sapiens ___________________________________________ 65
5. Synthèse __________________________________________________ 70
III LA FACE, LA MANDIBULE, LES DENTS ET LEUR EVOLUTION _77
1. La face ___________________________________________________ 77
1. Les os___________________________________________________ 77
3

2. Les muscles ______________________________________________ 78
2. La mandibule______________________________________________ 78
1. La mandibule _____________________________________________ 78
2. L’Articulation Temporo-Mandibulaire (ATM) ___________________ 79
3. Les rapports inter-maxillaires ________________________________ 81
3. Les dents__________________________________________________ 82
1. Morphologie______________________________________________ 82
2. Structure de l’émail ________________________________________ 84
3. Le système endodontique____________________________________ 85
CONCLUSION ________________________________________________ 86
REFERENCES BIBLIOGRAPHIQUES ____________________________ 87
LISTE DES ILLUSTRATIONS ___________________________________ 95
ANNEXE _____________________________________________________ 99
4

INTRODUCTION
L’évolution décrit les modifications des êtres vivants au cours du temps. Elle explique
la diversification de la vie, de ses premières formes jusqu'à l’ensemble des êtres vivants
actuels, par une chaîne de modifications buissonnantes.
Les dents jouent un rôle important pour comprendre l’évolution de notre lignée car :
• Elles se conservent bien dans les sols fossiles.
• Elles marquent l'appartenance du fossile à la famille à laquelle il appartient. Elles
témoignent donc du buissonnement des lignées.
• Elles conservent la trace du mode alimentaire et renseignent sur les conditions
écologiques. (54)
Nous allons voir dans un premier temps qu’elles ont été les différentes théories de
l’évolution établies jusqu’à aujourd’hui, théories de l’évolution en général mais aussi théories
concernant l’évolution de l’homme en particulier.
Dans une seconde partie, nous réaliserons un portrait plus ou moins détaillé de nos
lointains ancêtres, de Toumaï à Homo Sapiens, certains taxons décrits dans cette thèse ne sont
pas admis par l’ensemble de la communauté scientifique ; il nous a cependant semblé
intéressant de les faire apparaître ici de par leurs spécificités anatomiques.
Dans notre troisième partie nous ferons la synthèse des grands traits de l’évolution de
la face, de l’articulation temporo-mandibulaire et des dents en se basant sur les descriptions
précédentes. De plus nous essayerons de comprendre ce qui a entraîné ces évolutions.
5
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
1
/
101
100%