16/10/2015 GOBBI Amandine L3 CR : Julie Chapon SNP Pr

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Bases neurologiques de la vision, physiologie de la vision
16/10/2015
GOBBI Amandine L3
CR : Julie Chapon
SNP
Pr A. TREBUCHON-DA FONSECA
20 pages
Bases neurologiques de la vision, physiologie de la vision
A. Rappels sur les signaux visuels et la lumière
Les stimulus visuels sont des ondes électromagnétiques dont les longueurs d'ondes sont comprises entre 400
nm (violet) à 700 nm (rouge). Le spectre visible pour l'homme va donc du violet au rouge.
L'homme est capable de distinguer environ 200 nuances de couleurs. Ces nuances de couleurs dépendent du
niveau d'éclairement.
Une phrase connue est que « la nuit tous les chats sont gris ». En effet lorsque l'on se trouve dans la pénombre,
la vision des couleurs est quasi absente, voire nulle.
Ainsi pour pouvoir voir le spectre des couleurs il est nécessaire d'avoir un éclairement suffisant.
Analyse de l'image :
On observe des taches noires et blanches sur une
image : un dalmatien apparaît au premier coup d'œil
pour certain alors que pour d'autres le fait d'évoquer la
présence du dalmatien permet de le découvrir et de le
situer sur l'image alors que ce n'était pas du tout le cas
au premier abord : c'est l'effet Topdown ! Il s'agit de la
combinaison de l'information visuelle à différents
niveaux associée au savoir interne (Le dalmatien → il y
a du noir + du blanc → Je le vois!).
Cet effet est plus ou moins présent chez les individus.
Nos perceptions sont ainsi dépendantes de notre savoir
et une personne qui n'a jamais vu de dalmatien
auparavant ne peut pas le voir sur cette image.
CR : pour ceux qui ne le voient pas, le dalmatien se
trouve sur la droite de l'image, de biais tête baissée.
1/20
Plan :
A. Rappels sur les signaux visuels et la lumière
B. L'œil
I. Anatomie de l'œil
II. Formation de l'image sur la rétine
III. La rétine
C. Transfert à travers les voies optiques centrales (voie rétino-géniculo-corticale)
I. Anatomie
II. Organisation des voies optiques, notion de champ visuel
III. Corps genouillé latéral du thalamus
D. Traitement de l'information corticale
I. Cortex visuel primaire
II. Cortex visuel associatif

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Bases neurologiques de la vision, physiologie de la vision
Autre exemple :
On observe une illusion de 2 triangles par les 3 pacs man (un avec le trait noir et
un autre non dessiné mais que l'on peut deviner).
Ces 3 pacs-man sont orientés et pour pouvoir voir le triangle blanc (qui n'existe
pas en réalité) notre cerveau synchronise certaines parties de l'information visuelle
pour arriver à le voir.
Les messages lumineux transmis par la rétine passent ensuite par un relais au
niveau du thalamus pour au final rejoindre le cortex cérébral afin être traités, c'est ce qui va nous permettre
d'avoir une vision consciente permettant d'identifier et de localiser les objets.
B. L'œil
L'œil permet de transformer la lumière (onde électromagnétique) en un message nerveux (potentiel d'action,
PA) au niveau de la rétine, c'est la phototransduction.
I. Rappel : anatomie de l'œil
Avant d'arriver sur la rétine, la lumière passe par la cornée, le cristallin pour ensuite se focaliser sur un point
précis de la rétine au niveau du fond du globe oculaire là où sont concentrés l'ensemble des photorécepteurs et
le départ du nerf optique.
La cornée est une lentille convergente (puissance de réfraction de 42 dioptries) qui permet la focalisation de
l'image sur la rétine.
II. Formation de l'image sur la rétine
Trois éléments permettent la formation d'une image sur la rétine :
1) La cornée est une lentille convergente avec une puissance de réfraction de 42 dioptries permettant la
focalisation de l'image sur la rétine. Si la convexité cornéenne est anormale, l'image se formera soit en
avant (myopie) soit en arrière (hypermétropie) de la rétine.
2) L'iris va permettre une adaptation continue aux différentes intensités de lumière. Il est situé entre la
cornée et le cristallin et est un muscle lisse composé de deux types de fibres :
2/20

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Bases neurologiques de la vision, physiologie de la vision
- Lors d'une lumière ambiante sombre on observe une
dilatation de la pupille grâce aux fibres radiaires → on
parle de mydriase.
- En présence d'une forte luminosité, on observe un
contraction de la pupille grâce aux fibres circulaires →
on parle de myosis.
Si l'on est trop ébloui l'information se perd, et on a du mal à l'apprécier.
Les muscles de l'iris sont contrôlés par le système nerveux végétatif, non contrôlé de manière
consciente: le système orthosympathique assure la mydriase alors que le parasympathique permet le
myosis.
3) Le cristallin en ajoutant 12 dioptries, va permettre l'adaptation de l'image des objets proches (rôle dans
la vision de près). Il s'agit également d'une lentille convergente et à la différence de la cornée c'est un
système dynamique. Il y a un système de modification de courbure du cristallin grâce aux muscles
ciliaires : l’accommodation.
→ Lors de la presbytie la perte de plasticité du cristallin nécessite un besoin d'accommodation externe
avec une paire de lunettes pour voir de près. CR : la cataracte correspond à une opacification du
cristallin.
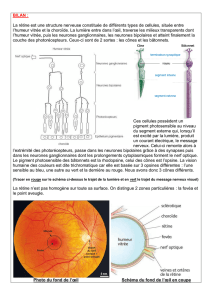
III. La rétine
La rétine est un tissu organisé qui permet de transformer la lumière en PA.
a. Anatomie de la rétine
Elle possède une organisation laminaire en 3 couches de cellules :
•Partie proximale (la + proche de la lumière) : cellules ganglionnaires. Les prolongements axonaux des
cellules ganglionnaires donnent le nerf optique.
3/20

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Bases neurologiques de la vision, physiologie de la vision
•Partie moyenne : cellules bipolaires
•Partie distale (partie externe de la rétine): cellules réceptrices, avec une couche de photorécepteurs (2
types de récepteurs : les cônes et les bâtonnets).
Ainsi avant d'atteindre la partie distale avec la présence des photorécepteurs la lumière traverse des couches
cellulaires qui ne vont pas contribuer à sa perception.
Les photorécepteurs ne sont pas répartis de manière égale sur toute la rétine.
Expérience : Lorsqu'on fixe un point précis (croix sur le tableau) dans une salle on a du mal à décrire les
caractéristiques de l'objet qui se situe sur le coté de notre champs visuel (couleur des bornes de sortie par
exemple). On teste ainsi notre vision centrale versus notre vision périphérique. La vision centrale est une vision
colorée, précise contrairement à la vision périphérique qui est plus une vision du mouvement et où la vision
colorée est beaucoup moins précise. Ce phénomène est du à la concentration quasi-nulle en cône au niveau de
la périphérie de la rétine.
On a donc un système hyper puissant et adapté d'orientation de la tête et des yeux, très connecté à la voie
visuelle, pour compenser ce phénomène et avoir une vision optimale.
La macula lutea, (tache jaune) = zone de la rétine caractérisée par une concentration maximale de cônes.
Située au fond de l'œil, dans l’axe de la pupille, la macula a un diamètre d’environ 2 mm.
La macula contient en son centre une petite dépression, la fovéa: entièrement composée de cônes serrés les uns
contre les autres, celle-ci est la zone d’acuité maximale de l'œil, c’est-à-dire celle qui donne la vision la plus
précise en éclairage diurne.
b. Les photorécepteurs
Il existe 2 types de photorécepteurs :
–les récepteurs cônes de forme cylindrique (à droite sur le schéma)
–et les récepteurs bâtonnets de forme plus rectangulaire (à gauche
sur le schéma)
Ces 2 types de cellules ne différent dans leur structure que par leur partie
terminale (partie externe) et se compose de 4 parties :
1) La partie externe avec les photorécepteurs présents dans la partie
externe de la rétine, ces derniers se composent d'un empilement de
disques (photopigments).
2) Le segment interne (contient le corps cellulaire et la terminaison
synaptique)
3) Le corps cellulaire
4) Et la terminaison synaptique permettant la libération de
neuromédiateur vers les cellules bipolaires.
4/20

SYSTÈME NEUROSENSORIEL ET PSYCHIATRIE – Bases neurologiques de la vision, physiologie de la vision
Les cônes sont surtout présents dans la partie centrale de la rétine au niveau d'une petite dépression appelé la
fovéa (centre de la tache jaune ou de la macula), tandis que l'on retrouve les bâtonnets plus dans la partie
latérale.
La proportion de cônes et de bâtonnets n'est pas identique. On a beaucoup plus de bâtonnets, 120 millions
contre 6 millions de cônes. Surtout retenir que l'on a 20 fois plus de bâtonnets, ce qui est énorme !!!
Les bâtonnets permettent la vision du contraste fin, de la luminosité, et du mouvement. Ils sont responsables de
la vision scotopique (ou vision de nuit) avec une forte sensibilité aux changements d'intensité lumineuse
(1000 fois plus sensible à la lumière que les cônes) et une faible vision des couleurs (la nuit on a du mal à
différencier les couleurs).
Les cônes sont responsable de la vision photopique (ou vision de jour).
Représentation des cônes et des bâtonnets sous la forme de leur perception des longueurs d'ondes
Les cônes ont la particularité d'avoir une réaction particulière à certaines longueurs d'ondes. Il existe trois types
de cônes avec des pigments différents, spécialisés dans la vision des couleurs sensibles à trois grand types de
longueurs d’onde (courte moyenne ou longue).
Représentation de la répartition spatiale des photorécepteurs sur la rétine :
Dans la partie centrale de la rétine on retrouve le
maximum de cônes et dans la partie latérale on
retrouve une proportion de bâtonnets importante
qui va décroitre au fur à mesure que l'on va aller
sur la périphérie de la rétine.
La vision précise est permise par l'oculomotricité et
par l'ensemble du système de la vision (cristallin,
cornée) qui va nous permettre de focaliser l'objet à
observer sur une petite partie de la rétine où se
concentrent les cônes. Les bâtonnets permettent
quant à eux d'élargir notre vision.
5/20
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%